
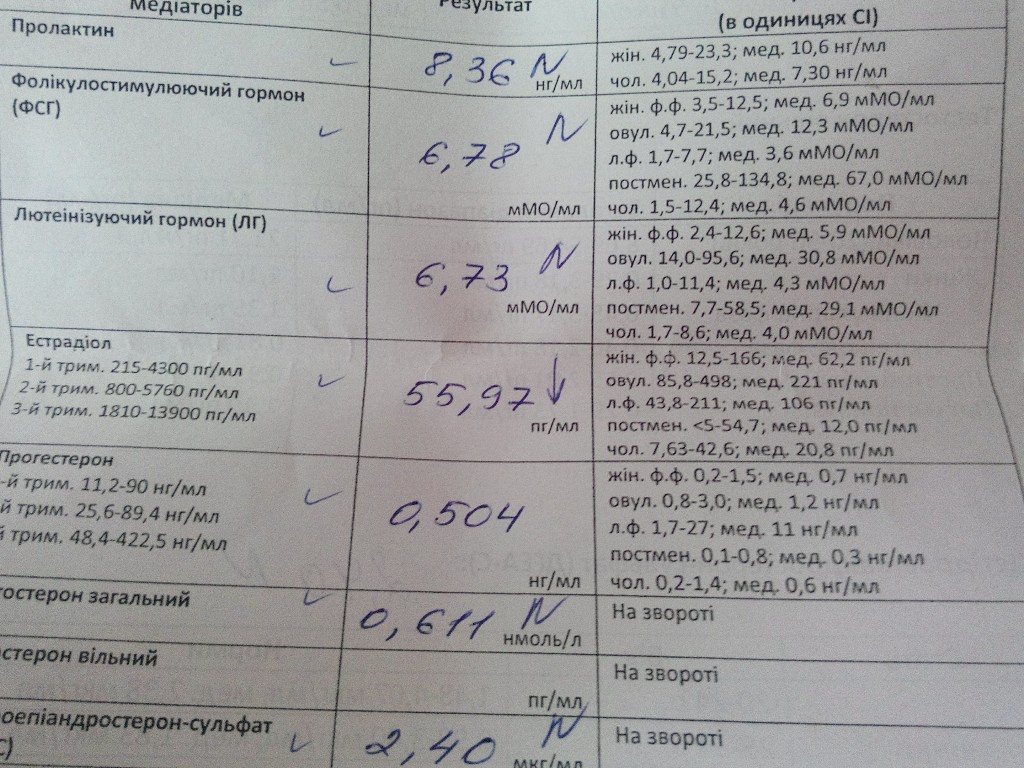
Функции пролактина: Значение пролактина в регуляции репродуктивной системы женщины
Значение пролактина в регуляции репродуктивной системы женщины
Среди многообразия причин приводящих к нарушению полноценной функции яичников и как результат – эндокринному бесплодию обусловленному отсутствием овуляции, особое место занимает феномен гиперпролактинемии.
Гиперпролактинемия – повышенное содержание пролактина (одного из важнейших гормонов гипофиза) в сыворотке крови.
Пролактин – это уникальный гормон, обладающий более чем тремястами различными биологическими эффектами. Основным местом синтеза пролактина является аденогипофиз – один из важнейших органов регуляции функции эндокринных желез, расположенный в головном мозге. Интересно отметить, что кроме гипофиза пролактин может синтезироваться и в других органах: в молочных и слезных железах, миометрии (мышцах матки), селезенке, коже, лимфатических узлах и костном мозге. Рецепторы, воспринимающие его сигнал, рассеяны практически по всему организму, включая органы иммунной системы. Спектр действия пролактина шире, чем суммарно у всех остальных гипофизарных гормонов.
Влияние пролактина на органы репродуктивной системы проявляется практически на всех уровнях регуляции. Так, еще в период полового созревания, пролактин обеспечивает процессы роста и структурного развития молочной железы. В яичниках, благодаря пролактину, происходит синхронизация процессов созревания фолликулов и овуляции. В матке пролактин повышает количество рецепторов к прогестерону и эстрогенам, благоприятно влияет на процесс имплантации эмбриона. Таким образом, недооценивать значение пролактина в регуляции репродуктивной системы невозможно. Однако основная роль этого уникального гормона – стимуляция синтеза молока железистой тканью молочных желез, проявляется после родов, когда имеется физиологическое повышение уровня пролактина в крови.
Таким образом, если в силу ряда причин (травмы и новообразования в головном мозге, прием определенных препаратов, период кормления грудью, стрессы и т.д.) уровень пролактина в сыворотке крови превышает физиологические значения, то это крайне негативно сказывается на репродуктивной функции.
Необходимо отметить, что сниженная функция щитовидной железы (гипотиреоз) так же может сопровождаться повышением уровня пролактина (тиреолиберин в повышенных количествах стимулирует синтез тиреотропного гормона и пролактина за счет перекрестного эффекта), поэтому крайне важно, при различных нарушениях меструального цикла наряду с пролактином, определять и уровень тиреотропного гормона (ТТГ) в сыворотке крови.
Повышение уровня пролактина может быть первичным, когда причина заключается в повышенной функции лактотрофов (клетки в гипофизе вырабатывающие пролактин) или в увеличении их количества, что бывает при опухолях гипофиза (пролактинома). Существуют и вторичные формы гиперпролактинемии связанные с различными заболеваниями и новообразованиями в головном мозге, приемом лекарственных препаратов, опосредованно стимулирующих образование пролактина (оральные контрацептивы, церукал, метилдопа, галаперидол и т.
Когда же следует задуматься о возможных нарушениях синтеза пролактина?
В первую очередь, при нерегулярном менструальном цикле. Наиболее часто повышение уровня пролактина в крови провоцирует задержки, а в более тяжелых случаях полное прекращение менструаций. Кроме того, если у женщины появляются выделения из молочных желез в виде молозива, усиливающиеся при надавливании, либо лакторрея (самопроизвольное выделение молока), следует срочно обратиться к специалисту. Минимальный перечень обследования должен включать в себя: определение содержания пролактина и ТТГ в крови, проведение ультразвукового (у женщин моложе 35 лет) или рентгенологического исследование молочных желез.
Следует обратить внимание, что даже если у женщины сохранен ритм менструаций, а беременность не наступает, необходимо, в первую очередь, определить содержание пролактина в сыворотке крови.
В зависимости от уровня пролактина врач принимает решение о целесообразности проведения МРТИ головного мозга, с целью исключения гормональноактивной опухоли гипофиза – пролактиномы. Как правило, микро (до 1 см) и макропролактиномы (более 1-го см) сопровождаются значительным увеличением содержания пролактина в крови (свыше 1500-2000 мМЕ/л). Менее значимые повышения – могут свидетельствовать о функциональной гиперпролактинемии.
Как правило, микро (до 1 см) и макропролактиномы (более 1-го см) сопровождаются значительным увеличением содержания пролактина в крови (свыше 1500-2000 мМЕ/л). Менее значимые повышения – могут свидетельствовать о функциональной гиперпролактинемии.
В этой связи, очень важно провести как минимум трехкратноеисследование гормона с промежутком в несколько дней и желательно, в одно и то же время.
Если диагноз – гиперпролактинемия оказывается подтвержденным, то прежде чем планировать беременность необходимо провести курс лекарственной терапии, направленной на нормализацию уровня пролактина. Длительность лечебного воздействия всегда индивидуальна, но, как правило, 3-6 месяцев стабильной компенсации оказывается достаточным. Если повышение пролактина обусловлено аденомой гипофиза, то необходимость в специфической терапии сохраняется на протяжении как периода подготовки к беременности, так и всего периода вынашивания и лактации.
Наконец, следует отметить наличие в клинической практике феномена «Макропролактинемии». Этот исключительно лабораторный феномен, обусловленный тем, что в крови у пациента определяют повышенные титры пролактина, а какая-либо клиническая симптоматика полностью отсутствует. Данное явление может быть вызвано тем, что повышение уровня гормона обусловлено биологически низко активным «big» и «big-big» пролактином с высокой молекулярной массой. В этом случае никакие лекарственные препараты принимать не нужно.
Этот исключительно лабораторный феномен, обусловленный тем, что в крови у пациента определяют повышенные титры пролактина, а какая-либо клиническая симптоматика полностью отсутствует. Данное явление может быть вызвано тем, что повышение уровня гормона обусловлено биологически низко активным «big» и «big-big» пролактином с высокой молекулярной массой. В этом случае никакие лекарственные препараты принимать не нужно.
Пролактин имеет довольно выраженный циркадный ритм с максимальным выбросом гормона во время ночного сна. Поэтому кровь для анализа берут утром, не ранее, чем через 3 часа после пробуждения. Нормативные значения пролактина в сыворотке крови у женщин составляют от 67 до 726 мМЕ/л;
Перед проведением исследования необходимо: за 1 день исключить половые контакты и тепловые воздействия (сауну), за 1 час — курение. Т.к. на уровень пролактина большое влияние оказывают стрессовые ситуации, желательно исключить факторы, влияющие на результаты исследований: физическое напряжение (бег, подъем по лестнице), эмоциональное возбуждение, бессонная ночь и т. п.
п.
Пролактин (S-Prol) – SYNLAB Eesti
Пролактин синтезируется в передней доле гипофиза. Его синтез и высвобождение подчиняются стимулирующему и ингибирующему действию гипоталамуса. Клинически важно то, что тиреотропин (TSH) и пролактин имеют один и тот же высвобождающий их гормон – TRH. Пролактин стимулирует функцию и рост молочных желез, вызывает лактацию. Пролактину характерно пульсирующее выделение. В связи с менструальным циклом его концентрация в крови мало изменяется. Во время беременности концентрация пролактина в крови увеличивается и достигает своего наибольшего значения во время родов. Поскольку пролактин ингибирует функцию яичников, то постоянное кормление грудью одновременно является естественной контрацепцией. Содержание пролактина снижается равномерно при наступлении менопаузы.
У мужчин функция пролактина точно неизвестна. Высокая концентрация пролактина тормозит продукцию тестостерона.
Показания:
У женщин
- Аменорея, олигоаменоррея, ановуляторный цикл, недостаточность corpus luteum, галакторрея, выяснение причин вирилизма
- Диагностика и контроль за лечением пролактинсекретирующих опухолей
- Заболевания гипоталамуса
У мужчин
- Нарушение либидо и потенции, гипогонадизм, выяснение причин галакторреи
Метод анализа: Хемилюминисцентный метод
Референтные значения:
| < 3 л | 163 – 1040 mIU/L |
| 3 – 9 л Ж | 63 – 420 mIU/L |
| > 9 л Ж | 96 – 770 mIU/L |
| Постменопауза, Ж | 38 – 430 mIU/L |
| 3 – 12 л M | 63 – 420 mIU/L |
| > 12 л M | 53 – 360 mIU/L |
Интерпретация результата:
Для выяснения причин стерильности у женщин рекомендуется определить, в первую очередь, пролактин. Клиническое значение имеют только повышенные значения пролактина, который способствуют торможению функций половых желез. Как результат этого возникают олигоаменоррея или аменорея (нарушения менструального цикла), при которых частым симптомом является галакторрея (истечение молока).
Клиническое значение имеют только повышенные значения пролактина, который способствуют торможению функций половых желез. Как результат этого возникают олигоаменоррея или аменорея (нарушения менструального цикла), при которых частым симптомом является галакторрея (истечение молока).
Гиперпролактинемия:
- Заболевания гипоталамуса – нарушения секреции или транспорта пролактинингибирующего фактора (PIF), компрессия ножки гипофиза опухолями
- Примарный гипотиреоз, с которым сопровождается увеличение содержания TRH
- Заболевания гипофиза – опухоли (пролактинома, другие аденомы), гиперплазия лактотропных клеток передней доли. Содержание общего пролактина >5000 mIU/L у небеременных указывает на пролактиному.
- Идиопатическая гиперпролактинемия
- Физиологические причины – беременность, кормление, сон, массаж груди, гипогликемия, физическая нагрузка и стресс. При беременности концентрация пролактина в 10-20 раз больше
- Лекарственные препараты – антидепрессанты, циметидин, эстрогены, антигипертензивные препараты, нейролептики
- Причиной гиперпролактинемии может быть также макропролактин, который не является биологически активным.
 Каждое четвертое повышенное значение пролактина вызвано макропролактином, поэтому до начала лечения гиперпролактинемии необходимо исключить наличие макропролактина.
Каждое четвертое повышенное значение пролактина вызвано макропролактином, поэтому до начала лечения гиперпролактинемии необходимо исключить наличие макропролактина.
 Каждое четвертое повышенное значение пролактина вызвано макропролактином, поэтому до начала лечения гиперпролактинемии необходимо исключить наличие макропролактина.
Каждое четвертое повышенное значение пролактина вызвано макропролактином, поэтому до начала лечения гиперпролактинемии необходимо исключить наличие макропролактина. Медицинский центр «Консультант» — Иммунохимические исследования (гормоны и онкомаркеры)
Пролактин (от англ. promotion – «стимулирование», lactation – «лактация») образуется в лактотрофах аденогипофиза. Основной функцией этого гормона является обеспечение лактации в молочных железах кормящей грудью женщины. Причем беременность и лактация являются единственными состояниями, при которых стойкое повышение уровня пролактина является нормой, в остальных случаях это нарушение.
Макропролактин — это пролактин, связанный в иммунные комплексы с аутоантителами. Являются ли эти аутоантитела непосредственной причиной заболевания или возникают в ответ на повышение концентрации пролактина до конца не ясно. В отличие от мономера пролактина, более крупный макропролактин удаляется почками дольше. Поэтому при макропролактинемии уровень пролактина часто значительно повышается, до 700мМе/л и более. При этом в результате взаимодействия с аутоантителом активность пролактина в молекуле макропролактина сильно снижена. Поэтому, в отличие от гиперпролактинемии, вызванной повышением концентрации пролактина в форме мономера (истинной гиперпролактинемии), макропролактинемия характеризуется бессимптомным течением или протекает в легкой форме. Самый распространенный ее признак – нарушение менструального цикла, а галакторея и бесплодие встречаются реже.
В отличие от мономера пролактина, более крупный макропролактин удаляется почками дольше. Поэтому при макропролактинемии уровень пролактина часто значительно повышается, до 700мМе/л и более. При этом в результате взаимодействия с аутоантителом активность пролактина в молекуле макропролактина сильно снижена. Поэтому, в отличие от гиперпролактинемии, вызванной повышением концентрации пролактина в форме мономера (истинной гиперпролактинемии), макропролактинемия характеризуется бессимптомным течением или протекает в легкой форме. Самый распространенный ее признак – нарушение менструального цикла, а галакторея и бесплодие встречаются реже.
Вот почему важно определять какой процент от пролактина составляет именно макропролактин.
Мономерный пролактин-один из наиболее важных гормонов, регулирующих репродуктивные функции человека .Он вырабатывается в передних долях гипофиза и наиболее важен для женщин. Кроме участия в обменных процессах и формировании вторичных половых признаков, основная функция этого гормона – осуществление лактации. Пролактин мономерный или по-другому – пост-ПЭГ – помогает в формировании молочных желез и стимулирует выработку молока после родов. Поэтому в период грудного вскармливание нужен повышенный уровень этого гормона. Кроме стимуляции лактации, он тормозит овуляцию и препятствует наступлению беременности.
Пролактин мономерный или по-другому – пост-ПЭГ – помогает в формировании молочных желез и стимулирует выработку молока после родов. Поэтому в период грудного вскармливание нужен повышенный уровень этого гормона. Кроме стимуляции лактации, он тормозит овуляцию и препятствует наступлению беременности.
Если мономерный пролактин повышен, женщина не может зачать ребенка. Этот гормон может вызвать не только исчезновение овуляции, но и вообще прекращение менструаций. Это приводит к бесплодию и многим заболеваниям женской половой сферы, поэтому анализ на его содержание чаще проводят гинекологи.
Причины повышения пролактина
Существует 2 вида гиперпролактинемиии — патологическая и физиологическая.
Патологическая возникает при некоторых заболеваниях:
- Болезни гипофиза (кисты мозга, аденома гипофиза ).
- Болезни гипоталамуса (туберкулез, герминома, радиоактивное облучение, саркоидоз).
- Другие заболевания (цирроз печени, хронический простатит, первичный гипотиреоз, эстрогенпродуцирующие опухоли, идиопатическая (не ясна причина) гиперпролактинемия).

Физиологическая — когда гормон пролактин повышается у здорового человека.
И бывает она в таких случаях:
- Кормление грудью (лактация).
- Гипогликемия (голод).
- Спортивные нагрузки (активность на спортивных площадках, а также выполнение работы, которая связана с поднятием тяжестей).
- Половой акт.
- Стресс.
- Беременность.
- Медицинские процедуры (длительные сеансы массажа области шеи, так как уровень пролактина контролируется нервными узлами, расположенными в шейном отделе).
- Стимуляция сосков при физическом напряжении или некачественным бельем. Выделения из сосков молочных желез посылают импульсы в кору головного мозга и этим провоцируют чрезмерное выделение пролактина.
 Выделения из сосков молочных желез посылают импульсы в кору головного мозга и этим провоцируют чрезмерное выделение пролактина.
Выделения из сосков молочных желез посылают импульсы в кору головного мозга и этим провоцируют чрезмерное выделение пролактина.Повышение пролактина также могут вызывать различные медикаменты: антидепрессанты, эстрогены, нейролептики, амфетамины, наркотические средства опиатного ряда.
Чем опасен повышенный пролактин?
Собственно повышения уровня гормона не несет в себе опасности для жизни. Если заболевание вызвано аденомой гипофиза, то с ростом образования в гипофизе могут нарастать симптомы сдавливания соседних сосудов отделов мозга.
Для женщин высокий уровень гормона является серьезным препятствием для зачатия и рождения ребенка. У мужчин пролактин в большом количестве может вызвать снижение половой функции и качества спермы, кроме этого может развиться гинекомастия, т. е. увеличение молочных желез.
Уровень пролактина 700мМЕ/л и более считается критерием необходимости проводить исследование на макропролактин, так как при такой концентрации пролактина наиболее часто встречается гипермакропролактинэмия.
В нашем медицинском центре при выполнении исследования на пролактин при его значении 700мМЕ/л и более, будет автоматически выполнен тест на определение макропролактина без дополнительной оплаты.
При значении пролактина менее 700мМЕ/л проводить определение макропролактина нецелесообразно, так как это не имеет диагностического значения.
Если желательно проведения теста на макропролактин, независимо от результата пролактина, можно оформить тест «Определение пролактина с макропролактином»
Пролактин (Prolactin) (+ дополнительный тест на макропролактин при результате пролактина выше 700 мЕд/л)
Исследуемый материал Сыворотка крови
Метод определения
Метод определения пролактина: твёрдофазный хемилюминесцентный иммуноанализ.
Тест на макропролактин – метод иммунопреципитации иммунных комплексов полиэтиленгликолем (ПЭГ).
Полипептидный гормон, стимулирующий пролиферацию молочной железы и секрецию молока.
Пролактин вырабатывается в передней доле гипофиза, небольшое количество синтезируется периферическими тканями. При беременности вырабатывается также в эндометрии. Во время беременности пролактин поддерживает существование жёлтого тела и выработку прогестерона, стимулирует рост и развитие молочных желёз и образование молока. Это один из гормонов, способствующих формированию полового поведения. Пролактин регулирует водно-солевой обмен, задерживая выделение воды и натрия почками, стимулирует всасывание кальция. В целом пролактин активирует анаболические процессы в организме. Среди других эффектов можно отметить стимуляцию роста волос. Пролактин оказывает также модулирующее воздействие на иммунную систему.
Суточная секреция пролактина имеет пульсирующий характер. Во время сна его уровень растет. После пробуждения концентрация пролактина резко уменьшается, достигая минимума в поздние утренние часы. После полудня уровень гормона нарастает. В отсутствие стресса, суточные колебания уровня находятся в пределах нормальных значений. Во время менструального цикла в лютеиновую фазу уровень пролактина выше, чем в фолликулиновую. С 8-й недели беременности уровень пролактина повышается, достигая пика к 20 — 25 неделе, затем снижается непосредственно перед родами и вновь увеличивается в период лактации.
После полудня уровень гормона нарастает. В отсутствие стресса, суточные колебания уровня находятся в пределах нормальных значений. Во время менструального цикла в лютеиновую фазу уровень пролактина выше, чем в фолликулиновую. С 8-й недели беременности уровень пролактина повышается, достигая пика к 20 — 25 неделе, затем снижается непосредственно перед родами и вновь увеличивается в период лактации.
Макропролактин – это пролактин, связанный в иммунные комплексы с антителами, присутствующий в крови в варьирующих количествах. Он выводится из крови медленней, чем мономерный пролактин и может накапливаться в высокой концентрации. Эта форма пролактина обладает меньшей биоактивностью, пациенты с высоким содержанием макропролактина могут не иметь классических симптомов, характерных для повышения концентрации пролактина.
Результаты данного исследования следует учитывать при трактовке повышенных значений показателя пролактина, расхождении результатов исследования с общей клинической картиной, отсутствии воспроизводимости при проведении исследований в разных лабораториях. Обращаем внимание на то, что выполнение исследования на макропролактин не увеличивает стоимость определения пролактина. Выявление возможного значимого присутствия макропролактина в пробах гиперпролактинемических пациентов необходимо для исключения диагностических ошибок, необходимости назначения ненужных биохимических и рентгенологических исследований, а также предотвращения неадекватной лекарственной терапии или хирургического вмешательства.
Пределы определения: 12,6 мЕд/л-172200 мЕд/л
Высокий уровень пролактина. Лечение в клинике Агада.
Уровень пролактина.
Гиперпролактинемия появляется при высоком уровне содержания гормона пролактина в крови. Это может быть как вариантом нормы, так и проявлением болезни. Пролактин вырабатывается гипофизом. Нормальные значения пролактина различаются у мужчин и женщин, они также отличны у людей разного возраста. Максимальный результат в анализах по этому показателю у женщин детородного возраста. После наступления климакса уровень пролактина снижается в 1,5–2 раза. Выделение пролактина в течение дня происходит циклически. Наиболее высокий уровень наблюдается ранним утром часы, а наименьший с 9 до 11 часов утра.
Пролактин вырабатывается гипофизом. Нормальные значения пролактина различаются у мужчин и женщин, они также отличны у людей разного возраста. Максимальный результат в анализах по этому показателю у женщин детородного возраста. После наступления климакса уровень пролактина снижается в 1,5–2 раза. Выделение пролактина в течение дня происходит циклически. Наиболее высокий уровень наблюдается ранним утром часы, а наименьший с 9 до 11 часов утра.
Действие пролактина на организм.
Биологическая функция пролактина заключается в поддержке деторождения, его главное действие заключается в становлении и регуляции лактации. Гормон участвует в формировании молочных желез, способствуя росту млечных протоков во время беременности и в подростковом возрасте. Пролактин стимулирует выработку грудного молока и способствует естественному вскармливанию детей. Воздействие пролактина на организм не ограничено молочными железами. Он оказывает влияние на поджелудочную и щитовидную железу, надпочечники, яичники. В надпочечниках пролактин увеличивает выработку адреналина, норадреналина, кортизола, андрогенов, альдостерона. Все эти гормоны помогают кормящей женщине легче переносить эмоциональные и физические перегрузки. Под действием этого гормона в поджелудочной железе возрастает функционирование клеток, производящих инсулин, благодаря чему поступающие в организм питательные вещества используются максимально полно. Воздействие пролактина на щитовидную железу позволяет уменьшить синтез кальцитонина, а значит, увеличить содержание минеральных веществ в грудном молоке. К пролактину особенно чувствительны яичники. Высокая концентрация гормона тормозит овуляцию и вызывает нарушение менструального цикла. Пролактин регулирует работу иммунной системы, позволяя плоду развиваться в матке без реакции отторжения. Пролактин в высоких дозах подавляет либидо, также он участвует в наступлении оргазма. Повышение содержания пролактина в организме происходит во время беременности и в период грудного вскармливания, это нужно для рождения и нормального развития ребенка.
В надпочечниках пролактин увеличивает выработку адреналина, норадреналина, кортизола, андрогенов, альдостерона. Все эти гормоны помогают кормящей женщине легче переносить эмоциональные и физические перегрузки. Под действием этого гормона в поджелудочной железе возрастает функционирование клеток, производящих инсулин, благодаря чему поступающие в организм питательные вещества используются максимально полно. Воздействие пролактина на щитовидную железу позволяет уменьшить синтез кальцитонина, а значит, увеличить содержание минеральных веществ в грудном молоке. К пролактину особенно чувствительны яичники. Высокая концентрация гормона тормозит овуляцию и вызывает нарушение менструального цикла. Пролактин регулирует работу иммунной системы, позволяя плоду развиваться в матке без реакции отторжения. Пролактин в высоких дозах подавляет либидо, также он участвует в наступлении оргазма. Повышение содержания пролактина в организме происходит во время беременности и в период грудного вскармливания, это нужно для рождения и нормального развития ребенка.
Причины повышения пролактина.
Причины гиперпролактинемии могут быть органическими при заболеваниях гипофиза или функциональными при заболеваниях других органов. Поражения гипофиза приводят к большому увеличению уровня пролактина. Причиной органической гиперпролактинемии чаще всего является микроаденома гипофиза, при этом его концентрация может превышать 2000–4000 мМЕ/л. Функциональная гиперпролактинемия может появиться как реакция на длительный и сильный стресс. Кратковременное повышение уровня пролактина, как правило, без клинических проявлений называется транзиторной гиперпролактинемией. Это состояние может быть связано с нарушением сна, половым актом или приемом пищи. В некоторых случаях врачи не могут установить причину повышения уровня пролактина.
Повышенный пролактин. Симптомы у мужчин и женщин.
Гиперпролактинемия может как минимально влиять на самочувствие человека, так и давать крайне тяжелые проявления. Самыми распространенными жалобами у мужчин с повышенным уровнем пролактина являются жалобы на снижение потенции.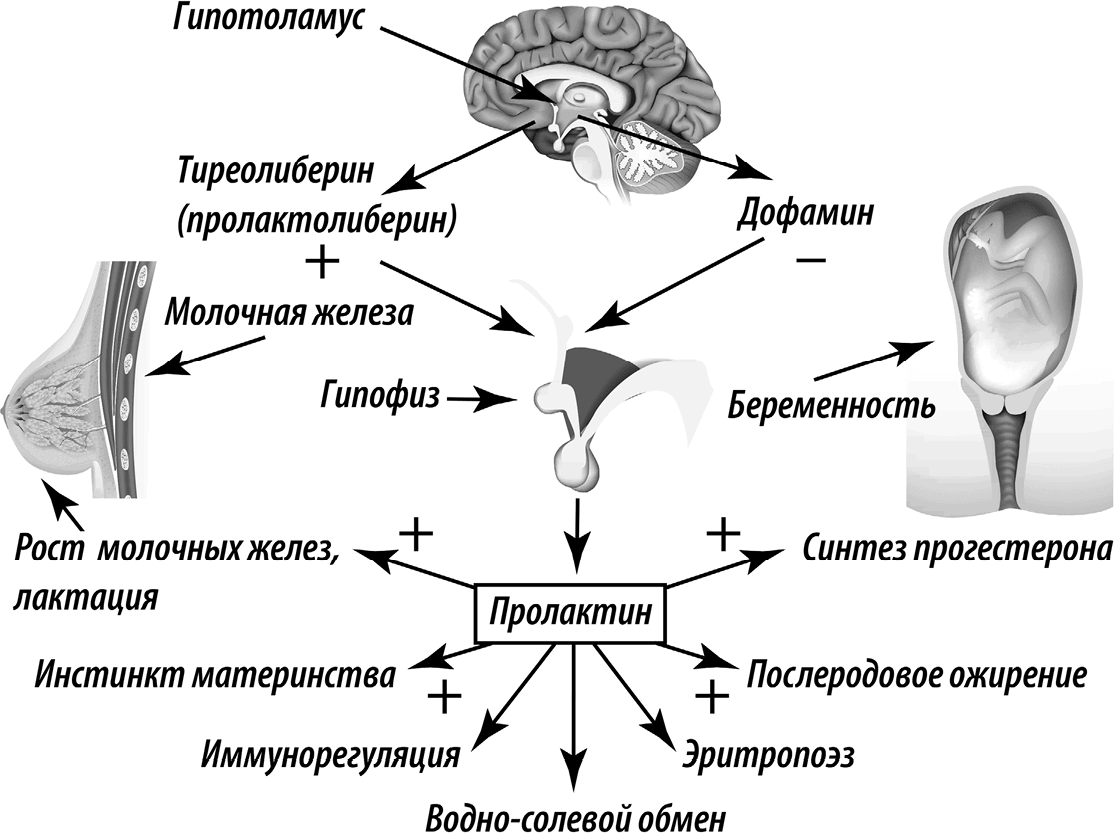 У женщин первым симптомом чаще всего являются жалобы на выделения из молочных желез. Симптомы гиперпролактинемии у женщин: отсутствие менструаций; нарушения менструального цикла, сухость влагалища, боль при половом акте, ановуляция и бесплодие. Симптомы гиперпролактинемии у мужчин: гинекомастия, уменьшение активности сперматозоидов, импотенция. Значительно чаще это заболевание встречается у женщин. Подтверждения диагноза у пациента проводится при помощи анализа на пролактин. При выявлении повышенного уровня, рекомендуется сдать гормоны щитовидной железы, анализ на сахар крови, пройти магнитно-резонансную или компьютерную томографию гипофиза.
У женщин первым симптомом чаще всего являются жалобы на выделения из молочных желез. Симптомы гиперпролактинемии у женщин: отсутствие менструаций; нарушения менструального цикла, сухость влагалища, боль при половом акте, ановуляция и бесплодие. Симптомы гиперпролактинемии у мужчин: гинекомастия, уменьшение активности сперматозоидов, импотенция. Значительно чаще это заболевание встречается у женщин. Подтверждения диагноза у пациента проводится при помощи анализа на пролактин. При выявлении повышенного уровня, рекомендуется сдать гормоны щитовидной железы, анализ на сахар крови, пройти магнитно-резонансную или компьютерную томографию гипофиза.
Обзор представлений о роли пролактина и пролактиновых рецепторов в репродуктивной системе женщин с наружным генитальным эндометриозом | Оразов
1. Ali S, Pellegrini I, Kelly PA. A prolactin-dependent immune cell line (Nb2) expresses a mutant form of prolactin receptor. J Biol Chem. 1991 Oct 25;266(30):20110-7.
J Biol Chem. 1991 Oct 25;266(30):20110-7.
2. Arden KC, Boutin JM, Djiane J, Kelly PA, Cavenee WK. The receptors for prolactin and growth hormone are localized in the same region of human chromosome 5. Cytogenet Cell Genet. 1990;53(2-3):161–5.
3. Bachelot A, Beaufaron J, Servel N, Kedzia C, Monget P, Kelly PA, Gibori G, Binart N. Prolactin independent rescue of mouse corpus luteum life span: identification of prolactin and luteinizing hormone target genes. Am J Physiol Endocrinol Metab. 2009 Sep;297(3):E676–84. doi: 10.1152/ajpendo.91020.2008.
4. Bazan JF. Structural design and molecular evolution of a cytokine receptor superfamily. Proc Natl Acad Sci USA. 1990 Sep;87(18):6934–8. doi: 10.1073/pnas.87.18.6934
5. Bernard V, Young J, Chanson P, Binart N. New insights in prolactin: pathological implications. Nature Reviews Endocrinology. 2015 Mar 17;11(5):265–75. doi: 10.1038/nrendo.2015.36
Bernard V, Young J, Chanson P, Binart N. New insights in prolactin: pathological implications. Nature Reviews Endocrinology. 2015 Mar 17;11(5):265–75. doi: 10.1038/nrendo.2015.36
6. Bernard V, Young J, Binart N. Prolactin — a pleiotropic factor in health and disease. Nature Reviews Endocrinology. 2019 Mar 21;15(6):356–65. doi: 10.1038/s41574-019-0194-6
7. Bernichtein S, Touraine P, Goffin V. New concepts in prolactin biology. J Endocrinol. 2010 Jul;206(1):1–11. doi: 10.1677/JOE-10-0069
8. Bethea CL, Neill JD. Lesions of the Suprachiasmatic Nuclei Abolish the Cervically Stimulated Prolactin Surges in the Rat*. Endocrinology. 1980 Jul;107(1):1–5. doi: 10.1210/endo-107-1-1
9. Bouilly J, Sonigo C, Auffret J, Gibori G, Binart N. Prolactin signaling mechanisms in ovary. Molecular and Cellular Endocrinology. 2012 Jun;356(1–2):80–7. doi: 10.1016/j.mce.2011.05.004
Bouilly J, Sonigo C, Auffret J, Gibori G, Binart N. Prolactin signaling mechanisms in ovary. Molecular and Cellular Endocrinology. 2012 Jun;356(1–2):80–7. doi: 10.1016/j.mce.2011.05.004
10. Boutin JM, Edery M, Shirota M, Jolicoeur C, Lesueur L, Ali S, Gould D, Djiane J, Kelly PA. Identification of a cDNA encoding a long form of prolactin receptor in human hepatoma and breast cancer cells. Mol Endocrinol. 1989 Sep;3(9):1455–61. doi: 10.1210/mend-3-9-1455
11. Brooks CL. Molecular Mechanisms of Prolactin and Its Receptor. Endocrine Reviews. 2012 Aug;33(4):504–25. doi: 10.1210/er.2011-1040
12. Brooks AJ, Waters MJ. The growth hormone receptor: mechanism of activation and clinical implications. Nature Reviews Endocrinology [Internet]. 2010 Sep 1 [cited 2020 Apr 24];6(9):515–525.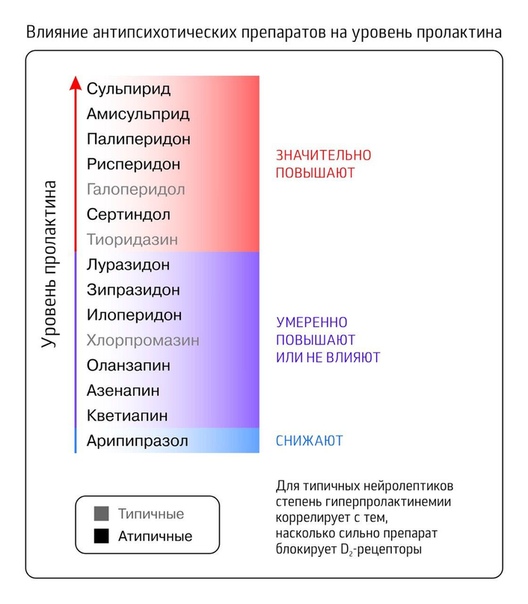 doi: 10.1038/nrendo.2010.123
doi: 10.1038/nrendo.2010.123
13. Cabrera-Reyes EA, Limon-Morales O, RiveroSegura NA, Camacho-Arroyo I, Cerbon M. Prolactin function and putative expression in the brain. Endocrine. 2017 Aug;57(2):199–213. doi: 10.1007/s12020-017-1346-x
14. Chen W, Guo X, Jin Z, Li R, Shen L, Li W1, Cai W. Transcriptional alterations of genes related to fertility decline in male rats induced by chronic sleep restriction. Syst Biol Reprod Med. 2020 Apr;66(2):99–111. doi: 10.1080/19396368.2019.1678694
15. Chen Y, Moutal A, Navratilova E, Kopruszinski C, Yue X, Ikegami M, et al. The prolactin receptor long isoform regulates nociceptor sensitization and opioid-induced hyperalgesia selectively in females. Science Translational Medicine. 2020 Feb 5;12(529):eaay7550. doi: 10. 1126/scitranslmed.aay7550
1126/scitranslmed.aay7550
16. Clevenger CV, Chang WP, Ngo W, Pasha TL, Montone KT, Tomaszewski JE. Expression of prolactin and prolactin receptor in human breast carcinoma. Evidence for an autocrine/paracrine loop. Am J Pathol. 1995;146(3):695–705.
17. Clevenger CV, Medaglia MV. The protein tyrosine kinase P59fyn is associated with prolactin (PRL) receptor and is activated by PRL stimulation of Tlymphocytes. Mol Endocrinol. 1994 Jun;8(6):674–81. doi: 10.1210/mend.8.6.7935483
18. Clevenger CV, Sillman AL, Hanley-Hyde J, Prystowsky MB. Requirement for prolactin during cell cycle regulated gene expression in cloned Tlymphocytes. Endocrinology. 1992 Jun;130(6):3216–22. doi: 10.1210/endo.130.6.1534539
19. Clevenger CV, Torigoe T, Reed JC. Prolactin induces rapid phosphorylation and activation of prolactin receptor-associated RAF-1 kinase in a Tcell line. J Biol Chem. 1994 Feb 25;269(8):5559–65.
Clevenger CV, Torigoe T, Reed JC. Prolactin induces rapid phosphorylation and activation of prolactin receptor-associated RAF-1 kinase in a Tcell line. J Biol Chem. 1994 Feb 25;269(8):5559–65.
20. Das R, Vonderhaar BK. Transduction of prolactin’s (PRL) growth signal through both long and short forms of the PRL receptor. Mol Endocrinol. 1995 Dec;9(12):1750–9. doi: 10.1210/mend.9.12.8614411
21. Esmaeilzadeh S, Mirabi P, Basirat Z, Zeinalzadeh M, Khafri S. Association between endometriosis and hyperprolactinemia in infertile women. Iran J Reprod Med. 2015;13(3):155–60.
22. Eyal O, Jomain JB, Kessler C, Goffin V, Handwerger S. Autocrine prolactin inhibits human uterine decidualization: a novel role for prolactin. Biol Reprod. 2007; 76:777e83. doi: 10.1095/biolreprod. 106.053058
106.053058
23. Gao Y, Shen M, Ma X, Li J, Wang B, Wang J, et al. Seven Hormonal Biomarkers for Diagnosing Endometriosis: Meta-Analysis and Adjusted Indirect Comparison of Diagnostic Test Accuracy. Journal of Minimally Invasive Gynecology. 2019 Sep;26(6):1026-1035.e4. doi: 10.1016/j.jmig.2019.04.004
24. Goffin V, Shiverick KT, Kelly PA, Martial JA. Sequence-Function Relationships Within the Expanding Family of Prolactin, Growth Hormone, Placental Lactogen, and Related Proteins in Mammals*. Endocrine Reviews. 1996 Aug;17(4):385–410. doi: 10.1210/edrv-17-4-385
25. Grattan DR, Kokay IC. Prolactin: a pleiotropic neuroendocrine hormone. J Neuroendocrinol. 2008 Jun;20(6):752–63. doi: 10.1111/j.1365-2826.2008.01736.x.
26. Grattan DR, Szawka RE. Kisspeptin and Prolactin. Semin Reprod Med. 2019 Mar;37(2):93–104. doi: 10.1055/s-0039-3400956.
Grattan DR, Szawka RE. Kisspeptin and Prolactin. Semin Reprod Med. 2019 Mar;37(2):93–104. doi: 10.1055/s-0039-3400956.
27. Gregoriou G, Bakas P, Vitoratos N, Papadias K, Goumas K, Chryssicopoulos A, et al. Evaluation of Serum ProlactinLevels in Patients with Endometriosisand Infertility. Gynecologic and Obstetric Investigation. 1999;48(1):48–51. doi: 10.1159/000010133
28. Grosdemouge I, Bachelot A, Lucas A, Baran N, Kelly PA, Binart N. Reproductive Biology and Endocrinology. 2003;1(1):12. doi: 10.1186/1477-7827-1-12
29. Huang C, Snider F, Cross JC. Prolactin Receptor Is Required for Normal Glucose Homeostasis and Modulation of β-Cell Mass during Pregnancy. Endocrinology. 2009 Apr;150(4):1618–26. doi: 10.1210/en.2008-1003
30. Bakowska JC, Morrell JI. The distribution of mRNA for the short form of the prolactin receptor in the forebrain of the female rat. Molecular Brain Research. 2003 Aug;116(1–2):50–8. doi: 10.1016/s0169-328x(03)00213-4
Bakowska JC, Morrell JI. The distribution of mRNA for the short form of the prolactin receptor in the forebrain of the female rat. Molecular Brain Research. 2003 Aug;116(1–2):50–8. doi: 10.1016/s0169-328x(03)00213-4
31. Kokay IC, Petersen SL, Grattan DR. Identification of prolactin-sensitive GABA and kisspeptin neurons in regions of the rat hypothalamus involved in the control of fertility. Endocrinology. 2011;152:526e35. doi: 10.1210/en.2010-0668
32. Kokay IC, Wyatt A, Phillipps HR, Aoki M, Ectors F, Boehm U, et al. Analysis of prolactin receptor expression in the murine brain using a novel prolactin receptor reporter mouse. Journal of Neuroendocrinology. 2018 Aug 23;30(9):e12634. doi: 10.1111/jne.12634
33. Ladyman SR, Hackwell ECR, Brown RSE. The role of prolactin in co-ordinating fertility and metabolic adaptations during reproduction. Neuropharmacology. 2020 May 1;167:107911. doi: 10.1016/j.neuropharm.2019.107911.
34. Lesueur L, Edery M, Ali S, Paly J, Kelly PA, Djiane J. Comparison of long and short forms of the prolactin receptor on prolactin-induced milk protein gene transcription. Proc Natl Acad Sci USA. 1991;88(3):824–828. doi: 10.1073/pnas.88.3.824
35. Li R, Kondegowda NG, Filipowska J, Hampton RF, Leblanc S, Garcia-Ocana A, et al. Lactogens Reduce Endoplasmic Reticulum Stress-induced Rodent and Human β-cell Death and Diabetes Incidence in Akita Mice. Diabetes. 2020 Apr 24;db190909. doi: 10.2337/db19-0909
36. Ling C, Svensson L, Oden B, Weijdegard B, Eden B, Eden S. Identification of functional prolactin (PRL) receptor gene expression: PRL inhibits lipoprotein lipase activity in human white adipose tissue. J Clin Endocrinol Metab. 2003;88:1804–8. doi: 10.1210/jc.2002-021137
37. Matalliotakis I, Panidis D, Vlassis G, Vavilis D, Neonaki M, Koumantakis E. PRL, TSH and their response to the TRH test in patients with endometriosis before, during, and after treatment with danazol. Gynecol Obstet Invest. 1996;42:183–86. doi: 10.1159/000291947
38. Mirabi P, Alamolhoda SH, Golsorkhtabaramiri M, Namdari M, Esmaeilzadeh S. Prolactin concentration in various stages of endometriosis in infertile women. JBRA Assist Reprod. 2019 Aug 22;23(3):225–9. doi: 10.5935/1518-0557.20190020
39. Motamedi B, Rafiee-Pour H-A, Khosravi M-R, Kefayat A, Baradaran A, Amjadi E, et al. Prolactin receptor expression as a novel prognostic biomarker for triple negative breast cancer patients. Annals of Diagnostic Pathology. 2020 Jun;46:151507. doi: 10.1016/j.anndiagpath.2020.151507
40. Nagano M, Kelly PA. Tissue distribution and regulation of rat prolactin receptor gene expression. Quantitative analysis by polymerase chain reaction. J Biol Chem. 1994 May 6;269(18):13337–45.
41. Ben-Jonathan N, LaPensee CR, LaPensee EW. What Can We Learn from Rodents about Prolactin in Humans? Endocrine Reviews. 2007 Dec 5;29(1):1–41. doi: 10.1210/er.2007-0017
42. Ben-Jonathan N, Mershon Jl, Allen Dl, Steinmetz RW. Extrapituitary Prolactin: Distribution, Regulation, Functions, and Clinical Aspects. Endocrine Reviews. 1996 Dec;17(6):639–69. doi: 10.1210/edrv-17-6-639
43. Ouhtit A, Morel G, Kelly PA. Visualization of Gene Expression of Short and Long Forms of Prolactin Receptor in Rat Reproductive Tissues. Biology of Reproduction. 1993 Sep 1;49(3):528–36. doi: 10.1095/biolreprod49.3.528
44. Panidis D, Vavilis D, Rousso D, Panidou E, Kalogeropoulos A. Provocative tests of prolactin before, during and after long-term danazol treatment in patients with endometriosis. Gynecological Endocrinology. 1992 Jan;6(1):19–24. doi: 10.3109/09513599209081002
45. Phillipps HR, Yip SH, Grattan DR. Patterns of prolactin secretion. Molecular and Cellular Endocrinology. 2020 Feb;502:110679. doi: 10.1016/j.mce.2019.110679
46. Qazi AM, Tsai-Morris C-H, Dufau ML. LigandIndependent Homo- and Heterodimerization of Human Prolactin Receptor Variants: Inhibitory Action of the Short Forms by Heterodimerization. Molecular Endocrinology. 2006 Aug;20(8):1912–23. doi: 10.1210/me.2005-0291
47. Rivera JC, Aranda J, Riesgo J, Nava G, Thebault S, López-Barrera F, et al. Expression and cellular localization of prolactin and the prolactin receptor in mammalian retina. Experimental Eye Research. 2008 Feb;86(2):314–21. doi: 10.1016/j.exer.2007.11.003
48. Sangeeta Devi Y, Halperin J. Reproductive actions of prolactin mediated through short and long receptor isoforms. Molecular and Cellular Endocrinology. 2014 Jan;382(1):400–10. doi: 10.1016/j.mce.2013.09.016
49. Shirota M, Banville D, Ali S, Jolicoeur C, Boutin J-M, Edery M, et al. Expression of Two Forms of Prolactin Receptor in Rat Ovary and Liver. Molecular Endocrinology. 1990 Aug;4(8):1136–43. doi: 10.1210/mend-4-8-1136
50. Silva JF, Henriques PC, Campideli-Santana AC, Araujo-Lopes R, Aquino NSS, Hipolito LTM, et al. Estradiol Potentiates But Is Not Essential for Prolactin-Induced Suppression of Luteinizing Hormone Pulses in Female Rats. Endocrinology. 2020 Feb 13;161(4). doi: 10.1210/endocr/bqaa022
51. Stevens AM, Yu-Lee LY. Multiple prolactinresponsive elements mediate G1 and S phase expression of the interferon regulatory factor-1 gene. Molecular Endocrinology. 1994 Mar;8(3):345–55. doi: 10.1210/mend.8.3.8015552
52. Thoreau E, Petridou B, Kelly PA, Djiane J, Mornon JP. Structural symmetry of the extracellular domain of the Cytokine/Growth hormone/Prolactin receptor family and Interferon receptors revealed by Hydrophobic Cluster Analysis. FEBS Letters. 1991 Apr 22;282(1):26–31. doi: 10.1016/0014-5793(91)80437-8
53. Toulis KA, Goulis DG, Kolibianakis EM, Venetis CA, Tarlatzis BC, Papadimas I. Risk of gestational diabetes mellitus in women with polycystic ovary syndrome: a systematic review and a metaanalysis. Fertility and Sterility. 2009 Aug;92(2):667–77. doi: 10.1016/j.fertnstert.2008.06.045
54. Tufa DM, Shank T, Yingst AM, Trahan GD, Shim S, Lake J, et al. Prolactin Acts on Myeloid Progenitors to Modulate SMAD7 Expression and Enhance Hematopoietic Stem Cell Differentiation into the NK Cell Lineage. Scientific Reports. 2020 Apr 14;10(1). doi: 10.1038/s41598-020-63346-4
55. Wang H, Gorpudolo N, Behr B. The Role of Prolactin– and Endometriosis-Associated Infertility. Obstetrical & Gynecological Survey. 2009 Aug;64(8):542–7. doi: 10.1097/OGX.0b013e3181ab5479
Пролактин (Prolaсtin)
Пролактин (Prolaсtin)
Пролактин – один из гормонов, выделяемых гипофизом. Пролактин необходим для нормального развития молочных желез, обеспечения лактации. Также он контролирует секрецию прогестерона и тормозит секрецию фолликулостимулирующего гормона, обеспечивая нормальный менструальный цикл. В крови мужчин и не беременных женщин он обычно присутствует в малых количествах. Уровень пролактина в крови обычно повышается во время беременности и в послеродовой период. У беременной женщины гормоны пролактина, эстрогена и прогестерона стимулируют выработку молока. Когда мать не кормит новорождённого грудью, пролактин в её крови падает обратно. Существует зависимость между тем, как часто ребенок сосёт грудь матери, и количеством пролактина и грудного молока, вырабатываемых её организмом. Другая причина повышения уровня пролактина – пролактинома – производящая пролактин опухоль гипофиза. Она встречается редко и, как правило, не бывает злокачественной. Наиболее часто такая опухоль развивается у женщин, но также бывает и у мужчин.
Подготовка к исследованию
Исследование проводится утром, натощак. С последнего приема пищи должно пройти не менее 8 часов. За 24 часа исключить половое сношение и тепловые воздействия (сауну), за 1 час курение. Желательно исключить факторы, влияющие на результаты исследований: физическое напряжение (бег, подъем по лестнице), эмоциональное возбуждение.
Показания к исследованию
— При симптомах пролактиномы (головные боли, ухудшение зрения, галакторея).
— При бесплодии и нарушении половой функции у мужчин и женщин.
— При пониженном уровне тестостерона у мужчин.
— Если у пациента выявлена пролактинома (для наблюдения за развитием опухоли).
— При подозрении на общую недостаточность функции гипофиза (в сочетании с тестом на гормон роста).
— Когда пациент принимает медикаменты, влияющие на выработку допамина организмом (в целях наблюдения за изменениями уровня пролактина).
Интерпретация
Референсные значения : еденица измерения мМЕ/л.
| Общая норма | Норма |
| для женщин | 40-530 |
| для мужчин | 53-360 |
Пол, возраст Референсные значения
1 — 12 мес Женский 4,2 — 633,9 мкМЕ/мл
Мужской 6,4 — 612,7 мкМЕ/мл
1 — 4 лет Женский 21,2 — 362,5 мкМЕ/мл
Мужской 48,8 — 279,8 мкМЕ/мл
4 — 7 лет Женский 33,9 — 277,7 мкМЕ/мл
Мужской 16,9 — 358,3 мкМЕ/мл
7 — 10 лет Женский 6,4 — 273,5 мкМЕ/мл
Мужской 40,3 — 245,9 мкМЕ/мл
10 — 13 лет Женский 40,3 — 203,5 мкМЕ/мл
Мужской 19,1 — 273,5 мкМЕ/мл
13 — 16 лет Женский 63,6 — 305,3 мкМЕ/мл
Мужской 33,9 — 351,9 мкМЕ/мл
16 — 18 лет Женский 44,5 — 390,1 мкМЕ/мл
Мужской 57,2 — 322,2 мкМЕ/мл
> 18 лет Женский (не беременные) 102 — 496 мкМЕ/мл
Мужской 86 — 324 мкМЕ/мл
Концентрация пролактина увеличивается при:
беременности и после родов, в период грудного вскармливания, нервно-психической анорексии, употреблении эстрогенов, трициклических антидепрессантов, опиатов, амфетаминов, препаратов, понижающих кровяное давление (резерпина, верапамила, метилдофы), заболеваниях, связанных с патологией гипоталамуса, гипотиреозе, заболеваниях почек, других онкологических заболеваниях гипофиза.
Воздействие стресса, возникшего в результате травмы, болезни или даже страха перед анализом, может приводить к умеренному увеличению уровня пролактина.
На результаты могут влиять
В течение суток уровень пролактина в крови меняется, увеличиваясь во время сна и достигая пиковых значений в утренние часы. Брать венозную кровь лучше после пробуждения пациента или хотя бы после того, как он отдохнул в спокойной обстановке минут 30.
Назначается в комплексе с
— лютеинизирующий гормон (ЛГ)
— фолликулостимулирующий гормон (ФСГ)
— тестостерон
— глобулин, связывающий половые гормоны (ГСПГ)
пролактин | физиология | Британника
Пролактин , также называемый лютеотропным гормоном (LTH) или лютеотропином , белковым гормоном, вырабатываемым гипофизом млекопитающих, который действует вместе с другими гормонами, чтобы инициировать секрецию молока молочными железами. В эволюционном масштабе пролактин — это древний гормон, выполняющий множество функций в обеспечении ухода за потомством (иногда называемый «родительским» гормоном). Это большая молекула белка, которая синтезируется и секретируется лактотрофами, которые составляют около 20 процентов передней доли гипофиза и расположены в основном в боковых областях железы.
Британская викторина
Человеческое тело
Возможно, вы знаете, что человеческий мозг состоит из двух половин, но какая часть человеческого тела состоит из крови? Проверьте обе половины своего разума в этой викторине по анатомии человека.
Физиологическое значение пролактина
У женщин основное действие пролактина — инициировать и поддерживать лактацию.У кормящих матерей тактильная стимуляция сосков и груди грудным младенцем блокирует секрецию гипоталамического дофамина (который обычно ингибирует пролактин) в гипофизарно-портальную циркуляцию гипофиза. Это приводит к резкому повышению концентрации пролактина в сыворотке крови, а затем к ее быстрому падению при прекращении кормления. Высокие концентрации пролактина в сыворотке подавляют секрецию гонадотропин-рилизинг-гормона (ГнРГ) гипоталамусом, тем самым снижая секрецию гонадотропинов (лютеинизирующий гормон и фолликулостимулирующий гормон), а также могут подавлять действие гонадотропинов на гонады.Таким образом, высокие концентрации пролактина в сыворотке крови во время кормления грудью снижают фертильность, защищая кормящих женщин от преждевременной беременности.
Поскольку пролактин поддерживает желтое тело яичника, которое является источником женского полового гормона прогестерона, он помогает поддерживать беременность. Кроме того, секреция пролактина во время беременности прогрессивно увеличивается. Секрецию пролактина также можно стимулировать высокими дозами эстрогенов, и она временно стимулируется стрессом и физическими упражнениями.Функция пролактина у мужчин неизвестна.
Регуляция секреции пролактина
Гипоталамическая регуляция секреции пролактина передней долей гипофиза отличается от гипоталамической регуляции других гормонов гипофиза в двух отношениях. Во-первых, гипоталамический контроль секреции пролактина является главным образом тормозящим, тогда как гипоталамический контроль секреции других гормонов передней доли гипофиза является стимулирующим. Таким образом, если передняя доля гипофиза отделена от влияния гипоталамуса, секреция пролактина увеличивается, тогда как секреция других гормонов передней доли гипофиза снижается.Гипоталамический фактор, который ингибирует секрецию пролактина, — это нейромедиатор дофамин, который не является нейропептидом, как и другие гипоталамические гормоны, регулирующие секрецию гормона передней доли гипофиза. Следовательно, препараты, имитирующие действие дофамина, полезны при лечении пациентов с высокими концентрациями пролактина в сыворотке крови.
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишитесь сейчасТакже существуют факторы, стимулирующие пролактин, и в их число входят гонадолиберин, тиреотропин-рилизинг-гормон и вазоактивный кишечный полипептид.Однако физиологическое значение этих факторов, стимулирующих пролактин, точно не определено. Одним из примеров фактора, стимулирующего пролактин, роль которого была определена, является эстроген, который стимулирует синтез и секрецию пролактина на поздних стадиях беременности, чтобы подготовить молочные железы к лактации.
Дефицит и избыток пролактина
Дефицит пролактина возникает в результате общего дефицита гормонов гипофиза, который характеризуется дефицитом других гормонов гипофиза в дополнение к пролактину.Основная причина дефицита гормона гипофиза — опухоль гипофиза. Наиболее ярким примером недостаточности пролактина является синдром Шихана, при котором передняя доля гипофиза беременных женщин частично или полностью разрушается во время или вскоре после родов. Этот синдром чаще встречается у женщин, у которых наблюдается обильное кровотечение во время родов. Больные женщины не производят грудное молоко и не могут кормить грудью своих младенцев. Дефицит пролактина не вызывает отклонений у женщин, которые не пытаются кормить грудью своих младенцев, и не вызывает отклонений у мужчин.
Повышенная секреция пролактина может быть вызвана повреждением ножки гипофиза, тем самым прерывая поток дофамина из гипоталамуса через гипофизарно-портальную циркуляцию к лактотрофам. Кроме того, повышенная секреция пролактина может быть вызвана пролактин-продуцирующими опухолями гипофиза, такими как лактотрофные аденомы или пролактиномы, а также некоторыми системными заболеваниями, особенно недостаточностью щитовидной железы. Многие лекарства, особенно те, которые используются для лечения психических или психических расстройств, высокого кровяного давления (гипертонии) и боли, также могут увеличивать секрецию пролактина.Однако у некоторых пациентов с высокими концентрациями пролактина в сыворотке (гиперпролактинемия) причина не видна, и, как говорят, у них наблюдается идиопатическая гиперпролактинемия.
У женщин репродуктивного возраста высокие концентрации пролактина в сыворотке крови приводят к снижению секреции гонадотропинов и, следовательно, к снижению циклической функции яичников. Частота менструальных циклов уменьшается (олигоменорея), и цикл может даже прекратиться (аменорея) вообще. Также наблюдаются симптомы дефицита эстрогена, такие как потеря полового влечения, сухость влагалища, бесплодие и, реже, нарушение лактации (галакторея).Высокие концентрации пролактина в сыворотке крови у женщин в постменопаузе обычно не связаны с какими-либо симптомами, хотя в очень редких случаях может возникнуть галакторея. У мужчин высокие концентрации пролактина в сыворотке также снижают секрецию гонадотропинов, но, следовательно, снижают функцию яичек, что приводит к низким концентрациям тестостерона в сыворотке. Основные симптомы — потеря полового влечения, эректильная дисфункция, мышечная слабость и бесплодие.
Пролактиномы — наиболее распространенный тип гормон-секретирующей опухоли гипофиза.У женщин они в четыре-пять раз чаще, чем у мужчин. Однако на момент постановки диагноза у мужчин пролактиномы, как правило, больше. Это различие объясняется тем, что нарушение менструального цикла является очень чувствительным индикатором избыточной секреции пролактина, тогда как снижение функции яичек у мужчин — нет. Пролактиномы часто вызывают головные боли, нарушения зрения, а также симптомы и признаки дефицита других гормонов гипофиза.
Большинство пациентов с пролактиномой лечатся препаратами, имитирующими действие дофамина, такими как бромокриптин и каберголин.Эти препараты приводят к быстрому снижению секреции пролактина и уменьшению размера опухоли. Однако в некоторых случаях препараты неэффективны или могут вызывать неприемлемые побочные эффекты, такие как тошнота, рвота и головные боли. Этих пациентов можно лечить хирургическим путем или лучевой терапией. Пациентам с небольшими симптомами — например, периодической задержкой менструального цикла — лечение может не потребоваться. У этих пациентов, как правило, наблюдаются нерастущие опухоли и умеренная гиперпролактинемия, которая не увеличивается.Препараты, подобные допамину, также снижают секрецию пролактина у пациентов с гиперпролактинемией от других причин, хотя предпочтительно устранить причину нарушения, если ее можно идентифицировать.
Роберт Д. УтигерУзнайте больше в этих связанных статьях Britannica:
Пролактин — обзор | ScienceDirect Topics
Действия пролактина на почки
Хотя существует четкое согласие, что рецепторы ПРЛ существуют в почках, в литературе мало единого мнения относительно потенциального действия пептида на почки.Это может быть результатом различных экспериментальных протоколов или использования препаратов гетерологичных пептидов. Отсутствие согласия также может быть результатом широко распространенного убеждения, что все эффекты ПРЛ в почках будут результатом пептида, доставляемого общим кровообращением. Это может быть не так, и уникальная изоформа ПРЛ может фактически быть эффективным агентом.
Horrobin et al. 17 вводили полуочищенный овечий ПРЛ внутримышечно пяти добровольцам-самцам. В дополнение к длительному недомоганию они наблюдали уменьшение объема мочи и экскреции натрия и калия с мочой, что сопровождалось увеличением натрия в плазме и осмоляльности.Волонтеры также испытывали чувство жажды. В то время нельзя было гарантировать абсолютную чистоту пептидного препарата, и, таким образом, нельзя было исключить загрязнение другими гормонами передней доли, а также нейрогипофизарными пептидами, окситоцином и вазопрессином. Это исследование на людях было задумано как отражение работы Локетта и др. № 20 , в котором антинатрийуретический, антидиуретический и антикалиуретический эффекты очищенных препаратов ПРЛ наблюдались у находящихся в сознании крыс в отсутствие очевидного воздействия на скорость клубочковой фильтрации.Таким образом, канальцевые эффекты для увеличения реабсорбции натрия были первыми предполагаемыми действиями ПРЛ.
Уменьшение объема мочи, наблюдаемое у людей-добровольцев, заставило многих предположить, что ПРЛ является уникальным антидиуретическим средством. Об этом также свидетельствует наблюдение на крысах, что инфузии овечьего ПРЛ снижают диурез и натрийурез при внутривенном введении физиологического раствора. 22 Однако в том же году появилось сообщение, в котором предполагалось, что антидиуретический эффект ПРЛ является результатом захвата рецепторов вазопрессина (АВП) в почках. 14 Действительно, Клапп и его коллеги элегантно показали, что пролактин, вводимый в мозг, или системно способствует высвобождению AVP и окситоцина. 35
В отсутствие AVP (крысы Brattleboro или крысы Sprague Dawley, нагруженные жидкостью для уменьшения эндогенной секреции AVP) рекомбинантный мышиный ПРЛ оказывает антинатриуетический и антидиуретический эффекты. Однако при наличии АВП действует мочегонное средство. Эти превосходные исследования показывают несколько важных недостатков более ранних работ: использование рекомбинантного материала, исключающего возможность контаминации другими реотропными факторами, использование видоспецифичного пептида и разделение потенциальных эффектов ПРЛ на действия АВП.Однако исследование 26 не изолирует проблему идентификации действия циркулирующего (гипофизарного происхождения) по сравнению с местно продуцируемым пептидом.
Он также не рассматривает возможность того, что пролактин почечного происхождения может оказывать действие на сегменты нефронов, отличные от тех, которые вводятся системно. Фактически, более недавние убедительные доказательства натрийуретического действия 12 в проксимальных извитых канальцах (ПКТ) опровергаются более ранней работой 6 , указывающей на антинатрийуретический эффект в мозговом веществе восходящей конечности (MAL) и дистальном извитом канальце (DCT).Действительно, хотя пролактин снижает активность Na +, K + -АТФазы при ПКТ, 19 , по-видимому, увеличивает активность при MAL и DCT. 6
Физиологическая значимость участка действия ПКТ пролактина, продуцируемого в почках, начинает получать поддержку. Здесь пролактин не только снижает активность Na +, K + -АТФазы, он сигнализирует об активации протеинкиназы A, протеинкиназы C и фосфорилировании клеточных белков (включая Na +, K + -ATPase) и через гетерологичное рекрутирование дофамина, D-1. рецептор. 12 Дофамин, действующий через рецептор D-1, является сильнодействующим натрийуретическим агентом и взаимодействует между допамином и другими натрийуретическими агентами (например, предсердным натрийуретическим фактором) в результате гетерологичного набора рецептора D-1. Crambert et al. 12 развивают свои аргументы еще на один шаг, показывая, что пролактин не может изменять активность Na +, K + -АТФазы у крыс SHR, линии, которая демонстрирует притупленный натрийуретический ответ на дофамин.
Пролактин — обзор | ScienceDirect Topics
Действия пролактина на почки
Хотя существует четкое согласие, что рецепторы ПРЛ существуют в почках, в литературе мало единого мнения относительно потенциального действия пептида на почки.Это может быть результатом различных экспериментальных протоколов или использования препаратов гетерологичных пептидов. Отсутствие согласия также может быть результатом широко распространенного убеждения, что все эффекты ПРЛ в почках будут результатом пептида, доставляемого общим кровообращением. Это может быть не так, и уникальная изоформа ПРЛ может фактически быть эффективным агентом.
Horrobin et al. 17 вводили полуочищенный овечий ПРЛ внутримышечно пяти добровольцам-самцам. В дополнение к длительному недомоганию они наблюдали уменьшение объема мочи и экскреции натрия и калия с мочой, что сопровождалось увеличением натрия в плазме и осмоляльности.Волонтеры также испытывали чувство жажды. В то время нельзя было гарантировать абсолютную чистоту пептидного препарата, и, таким образом, нельзя было исключить загрязнение другими гормонами передней доли, а также нейрогипофизарными пептидами, окситоцином и вазопрессином. Это исследование на людях было задумано как отражение работы Локетта и др. № 20 , в котором антинатрийуретический, антидиуретический и антикалиуретический эффекты очищенных препаратов ПРЛ наблюдались у находящихся в сознании крыс в отсутствие очевидного воздействия на скорость клубочковой фильтрации.Таким образом, канальцевые эффекты для увеличения реабсорбции натрия были первыми предполагаемыми действиями ПРЛ.
Уменьшение объема мочи, наблюдаемое у людей-добровольцев, заставило многих предположить, что ПРЛ является уникальным антидиуретическим средством. Об этом также свидетельствует наблюдение на крысах, что инфузии овечьего ПРЛ снижают диурез и натрийурез при внутривенном введении физиологического раствора. 22 Однако в том же году появилось сообщение, в котором предполагалось, что антидиуретический эффект ПРЛ является результатом захвата рецепторов вазопрессина (АВП) в почках. 14 Действительно, Клапп и его коллеги элегантно показали, что пролактин, вводимый в мозг, или системно способствует высвобождению AVP и окситоцина. 35
В отсутствие AVP (крысы Brattleboro или крысы Sprague Dawley, нагруженные жидкостью для уменьшения эндогенной секреции AVP) рекомбинантный мышиный ПРЛ оказывает антинатриуетический и антидиуретический эффекты. Однако при наличии АВП действует мочегонное средство. Эти превосходные исследования показывают несколько важных недостатков более ранних работ: использование рекомбинантного материала, исключающего возможность контаминации другими реотропными факторами, использование видоспецифичного пептида и разделение потенциальных эффектов ПРЛ на действия АВП.Однако исследование 26 не изолирует проблему идентификации действия циркулирующего (гипофизарного происхождения) по сравнению с местно продуцируемым пептидом.
Он также не рассматривает возможность того, что пролактин почечного происхождения может оказывать действие на сегменты нефронов, отличные от тех, которые вводятся системно. Фактически, более недавние убедительные доказательства натрийуретического действия 12 в проксимальных извитых канальцах (ПКТ) опровергаются более ранней работой 6 , указывающей на антинатрийуретический эффект в мозговом веществе восходящей конечности (MAL) и дистальном извитом канальце (DCT).Действительно, хотя пролактин снижает активность Na +, K + -АТФазы при ПКТ, 19 , по-видимому, увеличивает активность при MAL и DCT. 6
Физиологическая значимость участка действия ПКТ пролактина, продуцируемого в почках, начинает получать поддержку. Здесь пролактин не только снижает активность Na +, K + -АТФазы, он сигнализирует об активации протеинкиназы A, протеинкиназы C и фосфорилировании клеточных белков (включая Na +, K + -ATPase) и через гетерологичное рекрутирование дофамина, D-1. рецептор. 12 Дофамин, действующий через рецептор D-1, является сильнодействующим натрийуретическим агентом и взаимодействует между допамином и другими натрийуретическими агентами (например, предсердным натрийуретическим фактором) в результате гетерологичного набора рецептора D-1. Crambert et al. 12 развивают свои аргументы еще на один шаг, показывая, что пролактин не может изменять активность Na +, K + -АТФазы у крыс SHR, линии, которая демонстрирует притупленный натрийуретический ответ на дофамин.
Функция пролактина и предполагаемая экспрессия в головном мозге
А. Игначак, М. Каштельник, Т. Слива, Р.А. Корбут, К. Райда, Т.Дж. Гузик, Пролактин — не только лактотропин, это «новый» взгляд на «старый» гормон. J. Physiol. Pharmacol. 63 (5), 435–443 (2012)
CAS PubMed Google Scholar
R.J. Марано, Н. Бен-Джонатан, Мини-обзор: внегипофизарный пролактин: обновленная информация о распределении, регуляции и функциях. Мол. Эндокринол. 28 (5), 622–633 (2014)
PubMed PubMed Central Статья CAS Google Scholar
D.R. Граттан, И. Кокай, Пролактин: плейотропный нейроэндокринный гормон. J. Neuroendocrinol. 20 (6), 752–763 (2008)
CAS PubMed Статья Google Scholar
Р., Бейтс, Э., Лар, О., Риддл, Грубое действие пролактина и фолликулостимулирующего гормона на зрелый яичник и половые органы птицы. Являюсь. J. Physiol. I (II), 361–368 (1935)
N. Ben-jonathan, J.Л. Мершонф, Д.Л. Аллен, Р.В. Стейнмец, Внегипофизарный пролактин: распределение, регуляция, функции и клинические аспекты. Endocr. Ред. 17 (6), 639–669 (1996)
CAS PubMed Google Scholar
С. Харви, Внегипофизарный гормон роста. Эндокринная 38 (3), 335–359 (2010)
CAS PubMed Статья Google Scholar
С. Харви, К. Арамбуро, Э.Дж. Сандерс, Внегипофизарная продукция гормонов передней доли гипофиза: обзор. Эндокринная 41 (1), 19–30 (2012)
CAS PubMed Статья Google Scholar
Р.Дж., Уолш, Ф.Дж., Слаби, Б.И. Познер, Опосредованный рецептором механизм транспорта пролактина из крови в спинномозговую жидкость. Эндокринология 120 (5), 1846–1850 (1987)
R.S.E. Браун, А. Wyatt, R.E. Гербисон, П.Дж. Ноулз, С. Ladyman, N. Binart, W.A. Banks, D.R. Граттан, Транспорт пролактина в мозг мыши не зависит от рецептора пролактина. FASEB J. 30 (2), 1002–1010 (2016)
CAS PubMed Статья Google Scholar
Н. Эмануэле, Дж. Юргенс, М. Халлоран, Дж. Тентлер, А. Лоуренс, М. Келли, Ген пролактина крысы экспрессируется в ткани мозга: обнаружение нормальной и альтернативно сплайсированной матричной РНК пролактина.Мол. Эндокринол. 6 (1), 35–42 (1992)
CAS PubMed Google Scholar
К. Филдс, Э. Кулиг, Р. Ллойд, Обнаружение матричной РНК пролактина в молочных железах и других нормальных и опухолевых тканях с помощью полимеразной цепной реакции. Лаборатория. Вкладывать деньги. 68 (3), 354–360 (1993)
CAS PubMed Google Scholar
Х. Эманн, К. Зальциг, П.Ланг, Э. Фриауф, Х. Нотванг, Минимальные половые различия в экспрессии генов в верхнем оливковом комплексе крысы. Слышать. Res. 245 (1–2), 65–72 (2008)
PubMed Статья Google Scholar
W. DeVito, C. Avakian, S. Stone, C. Ace, Эстрадиол увеличивает синтез пролактина и рибонуклеиновую кислоту-мессенджер пролактина в выбранных областях мозга гипофизэктомированной самки крысы. Эндокринология 131 (5), 2154–2160 (1992)
CAS PubMed Статья Google Scholar
C.E. Roselli, S. Bocklandt, H.L. Stadelman, T. Wadsworth, E. Vilain, F. Stormshak, Экспрессия пролактина в мозге овец. Нейроэндокринология 87 (4), 206–215 (2008)
CAS PubMed Статья Google Scholar
В. ДеВито, Распределение иммунореактивного пролактина в головном мозге самцов и самок крыс: эффекты гипофизэктомии и внутрижелудочкового введения колхицина. Нейроэндокринология 47 (4), 284–289 (1988)
CAS PubMed Статья Google Scholar
В. Гоффин, П. Келли, Семейство рецепторов пролактина / гормона роста: взаимосвязь между структурой и функцией. J. Mammary Gland Biol. Неоплазия 2 (1), 7–17 (1997)
CAS PubMed Статья Google Scholar
С. Боле-Фейсот, В. Гоффин, М. Эдери, Н. Бинарт, П. Келли, Пролактин (ПРЛ) и его рецептор: пути передачи сигнала действия и фенотипы, наблюдаемые у мышей с нокаутом рецептора ПРЛ. Endocr. Ред. 19 (3), 225–268 (1998)
CAS PubMed Статья Google Scholar
М. Фриман, Б. Каньицка, А. Ленарт, Г. Надь, Пролактин: структура, функция и регуляция секреции. Physiol. Ред. 80 , 1523–1631 (2000)
CAS PubMed Google Scholar
К. Клевенджер, Дж. Клайн, Передача сигнала рецептора пролактина. Волчанка 10 (10), 706–718 (2001)
CAS PubMed Статья Google Scholar
X. Pi, J.L. Voogt, Половые различия и эстральный цикл: экспрессия мРНК рецептора пролактина в головном мозге крысы. Мол. Brain Res. 103 (1–2), 130–139 (2002)
CAS PubMed Статья Google Scholar
Дж. Баковска, Дж. Моррелл, Распределение мРНК для короткой формы рецептора пролактина в переднем мозге самки крысы. Brain Res. 116 (1–2), 50–58 (2003)
CAS Статья Google Scholar
J.C. Bakowska, J.I. Моррелл, Атлас нейронов, экспрессирующих мРНК длинной формы рецептора пролактина в переднем мозге самки крысы. J. Comp. Neurol. 386 (2), 161–177 (1997)
CAS PubMed Статья Google Scholar
X.J. Пи, Д. Граттан, Повышенная экспрессия коротких и длинных форм мРНК рецептора пролактина в ядрах гипоталамуса кормящих крыс. J. Mol. Эндокринол. 23 (1), 13–22 (1999)
CAS PubMed Статья Google Scholar
Т. Фудзикава, Х. Соя, Х. Йошизато, К. Сакагути, К. Дох-Ура, М. Танака, К. Накашима, Сдерживающий стресс усиливает экспрессию гена длинной формы рецептора пролактина в сосудистой оболочке сплетение. Эндокринология 136 , 5608–5613 (1995)
CAS PubMed Статья Google Scholar
Дж. Хираи, М. Нишита, Н. Накао, Т.Р. Сайто, М. Танака, Регулирование экспрессии гена рецептора пролактина в сосудистом сплетении крысы посредством транскрипционной активации нескольких первых экзонов во время постнатального развития и лактации. Exp. Anim. 62 (1), 49–56 (2013)
CAS PubMed Статья Google Scholar
S. Chiu, P.M. Мудрый, локализация мРНК рецептора пролактина в гипоталамусе путем гибридизации in situ.J. Neuroendocrinol. 6 (2), 191–199 (1994)
CAS PubMed Статья Google Scholar
Р.С. Браун, И. Кокай, А.Э. Хербисон, Д. Граттан, Распределение пролактин-чувствительных нейронов в переднем мозге мышей. J. Comp. Neurol. 518 (1), 92–102 (2010)
CAS PubMed Статья Google Scholar
Т.А. Мёдершайм, Т.Горба, П. Патипати, И. Кокай, Д. Grattan, C.E. Williams, A. Scheepens, Пролактин участвует в глиальных реакциях после очагового повреждения головного мозга молодых крыс. Неврология 145 (3), 963–973 (2007)
PubMed Статья CAS Google Scholar
L. Torner, S. Karg, A. Blume, M. Kandasamy, H.G. Kuhn, J. Winkler, L. Aigner, I.D. Neumann, пролактин предотвращает вызванное хроническим стрессом снижение нейрогенеза в гиппокампе взрослых и способствует судьбе нейронов.J. Neurosci. 29 (6), 1826–1833 (2009)
CAS PubMed Статья Google Scholar
E.A. Кабрера-Рейес, Э. Вергара-Кастаньеда, Н. Риверо-Сегура, М. Цербон, Половые различия в пролактине и его рецепторной экспрессии в гипофизе, гипоталамусе и гиппокампе крысы. Преподобный Мекс. Эндокринол. Метаб. Nutr. 2 (2), 60–67 (2015)
Google Scholar
X.J. Пи, Д. Граттан, Распределение иммунореактивности рецепторов пролактина в головном мозге крыс после овариэктомии, получавших эстроген. J. Comp. 394 , 462–474 (1998)
CAS Google Scholar
Институт мозговых исследований Аллена, Атлас мозга Аллена Мыши. http://mouse.brain-map.org/experiment/show/72340223 (2004 г.).
М. Шамгочян, С. Авакян, Н. Труонг, С. Стоун, К. Танг, В. Де Вито, Регулирование экспрессии рецептора пролактина эстрадиолом в головном мозге самок крыс.Нейроотчет. 6 (18), 2537–2541 (1995)
CAS PubMed Статья Google Scholar
П. Манн, Р. Бриджес, Экспрессия гена рецептора пролактина в переднем мозге беременных и кормящих крыс. Brain Res. Мол. Brain Res. 105 (1–2), 136–145 (2002)
CAS PubMed Статья Google Scholar
S. Chiu, R.D. Koos, P.M. Wise, Обнаружение мРНК рецептора пролактина (PRL-R) в гипоталамусе и гипофизе крыс. Эндокринология 130 (3), 1747–1749 (1992)
CAS PubMed Google Scholar
S.H. Ип, Р. Егучи, Д. Граттан, С.Дж. Банн, передача сигналов пролактина в гипоталамусе мыши в основном опосредуется преобразователем сигнала и активатором фактора транскрипции 5b, но не 5a. J. Neuroendocrinol. 24 (12), 1484–1491 (2012)
CAS PubMed Статья Google Scholar
Р. Дас, Б. Вондерхаар, Передача сигнала роста пролактина (ПРЛ) через длинные и короткие формы рецептора ПРЛ. Мол. Эндокринол. 9 , 1750–1759 (1995)
CAS PubMed Google Scholar
Дж. Харрис, П.М. Стэнфорд, С. Оукс, К.Дж. Орманди, Пролактин и рецептор пролактина: новые мишени старого гормона. Аня. Med. 36 (6), 414–425 (2004)
CAS PubMed Статья Google Scholar
Л. Торнер, Э. Тинахеро, Н. Ладжуд, А. Кинтанар-Стефано, Э. Ольвера-Кортес, Гиперпролактинемия ухудшает распознавание объектов без изменения пространственного обучения у самцов крыс. Behav. Brain Res. 252 , 32–39 (2013)
CAS PubMed Статья Google Scholar
М.Дж. Патил, М.А. Генри, А.Н. Акопян, Рецептор пролактина в регуляции возбудимости нейронов и каналов. Каналы 8 (3), 193–202 (2014)
PubMed PubMed Central Статья Google Scholar
Ф. Драго, Пролактин и сексуальное поведение: обзор. Neurosci. Biobehav. Ред. 8 , 433–439 (1984)
CAS PubMed Статья Google Scholar
D. Tejadilla, M. Cerbón, T. Morales, Prolactin снижает повреждающие эффекты эксайтотоксичности в дорсальном гиппокампе самки крысы независимо от гормонов яичников. Неврология 169 (3), 1178–1185 (2010)
CAS PubMed Статья Google Scholar
А. Ванойе-Карло, Т. Моралес, Э. Рамос, А. Мендоса-Родригес, М. Цербон, Нейропротективные эффекты лактации против лечения каиновой кислотой в спинном гиппокампе крысы. Horm. Behav. 53 (1), 112–123 (2008)
CAS PubMed Статья Google Scholar
В. Кабрера, Д. Канту, Э. Рамос, А. Ванойе-Карло, М. Цербон, Т. Моралес, Лактация является естественной моделью нейрозащиты гиппокампа от эксайтотоксичности.Neurosci. Lett. 461 (2), 136–139 (2009)
CAS PubMed Статья Google Scholar
В. Кабрера, Э. Рамос, А. Гонсалес-Аренас, М. Цербон, И. Камачо-Арройо, Т. Моралес, Лактация снижает активацию глии, вызванную эксайтотоксичностью в гиппокампе крыс. J. Neuroendocrinol. 25 (6), 519–527 (2013)
CAS PubMed Статья Google Scholar
Р. Браун, А. Гербисон, Д. Граттан, Влияние пролактина и лактации на дофаминовые нейроны A15 в ростральной преоптической области самок мышей. J. Neuroendocrinol. 27 (9), 708–717 (2015)
CAS PubMed Статья Google Scholar
J.A. Сегги, Г. Браун, Паттерны стрессовой реакции кортикостерона, пролактина и гормона роста в плазме у крыс после обработки или воздействия новой окружающей среды.Может. J. Physiol. Pharmacol. 53 (4), 629–637 (1975)
CAS PubMed Статья Google Scholar
A.C. de Moura, V.M. Лаццари, Р.О. Беккер, М. Гил, К.А. Рутшиллинг, Дж. Агнес, С. Алмейда, А. да Вейга, А. Лусион, М. Джовенардо, Экспрессия генов в ЦНС кормящих крыс с различными паттернами материнского поведения. Neurosci. Res. 99 , 8–15 (2015)
PubMed Статья CAS Google Scholar
C.M. Ларсен, И. Кокай, Д. Граттан, мужские феромоны инициируют индуцированный пролактином нейрогенез и улучшают материнское поведение у самок мышей. Horm. Behav. 53 (4), 509–517 (2008)
CAS PubMed Статья Google Scholar
Ф. Леви, М. Келлер, П. Пуиндрон, Обонятельная регуляция материнского поведения у млекопитающих. Horm. Behav. 46 (3), 284–302 (2004)
PubMed Статья Google Scholar
Х. Салаис-Лопес, Э. Лануса, К. Агустин-Павон, Ф. Мартинес-Гарсия, Настройка мозга для материнства: пролактин-подобная центральная передача сигналов у девственных, беременных и кормящих самок мышей. Структура мозга. Funct. 222 (2), 895–921 (2017)
PubMed Статья CAS Google Scholar
М. Фримарк, П. Дрисколл, Дж. Эндрюс, П.А. Келли, М. Ройстер, Онтогенез экспрессии гена рецептора пролактина в обонятельной системе крыс: потенциальные роли лактогенных гормонов в развитии обоняния.Эндокринология 137 (3), 934–942 (1996)
CAS PubMed Статья Google Scholar
T. Shingo, C. Gregg, E. Enwere, H. Fujikawa, R. Hassam, C. Geary, J.C. Cross, S. Weiss, Нейрогенез, стимулируемый беременностью в переднем мозге взрослой женщины, опосредованный пролактином. Наука 299 (5603), 117–120 (2003)
CAS PubMed Статья Google Scholar
Р. Бриджес, Д. Граттан, Пролактин-индуцированный нейрогенез в материнском мозге. Тенденции Эндокринол. Метаб. 14 (5), 199–201 (2003)
CAS PubMed Статья Google Scholar
N., Lajud, R., Gonzalez-Zapien, A., Roque, E., Tinajero, JJ, Valdez, C., Clapp, L., Torner, Введение пролактина в ранний постнатальный период жизни снижает гиппокампальный и нейрогенез обонятельной луковицы и приводит к депрессивному поведению во взрослом возрасте.Horm. Behav. 64 (5), 781–789 (2013)
А. Флеминг, М. Корсмит, Пластичность в материнской цепи: влияние материнского опыта на Fos-Lir в гипоталамических, лимбических и корковых структурах в послеродовая крыса. Behav. Neurosci. 110 (3), 567–582 (1996)
CAS PubMed Статья Google Scholar
Эляда Ю., Мизрахи А. Становление пластичности материнской цепи, лежащей в основе материнского поведения.Curr. Opin. Neurobiol. 35 , 49–56 (2015)
CAS PubMed Статья Google Scholar
J.N. Lundström, A. Mathe, B. Schaal, J. Frasnelli, K. Nitzsche, J. Gerber, T. Hummel, Материнский статус регулирует корковые реакции на запах тела новорожденных. Фронт. Psychol. 4 , 1–6 (2013)
Артикул Google Scholar
L.Коэн, А. Мизрахи, Пластичность во время материнства: изменения в нейронах возбуждающего и тормозного слоя 2/3 в слуховой коре. J. Neurosci. 35 (4), 1806–1815 (2015)
CAS PubMed Статья Google Scholar
К. Грегг, В. Шикар, П. Ларсен, Г. Мак, А. Хойнацки, В.В. Йонг, С. Вайс, Пластичность белого вещества и усиление ремиелинизации в материнской ЦНС. J. Neurosci. 27 (8), 1812–1823 (2007)
CAS PubMed Статья Google Scholar
М. Маэ, Э. Акбари, А. Флеминг, Число каллозальных олигодендроцитов у послеродовых крыс Sprague-Dawley. Brain Res. 24 (1267), 18–24 (2009)
Артикул CAS Google Scholar
П. Манн, Р. Бриджес, Лактогенная гормональная регуляция материнского поведения. Прог. Brain Res. 133 , 251–262 (2001)
CAS PubMed Статья Google Scholar
М. Нуман, М. Нуман, анализ поражения и нейроанатомического тракта с отслеживанием роли ядра ложа терминальной полоски в поведении при поиске и других аспектах материнской реакции у крыс. Dev. Psycobiol. 29 (1), 23–51 (1996)
CAS Статья Google Scholar
М. Нуман, Анализ нейронных схем материнского поведения у крысы. Acta Paediatr. Дополнение 397 , 19–28 (1994)
CAS PubMed Статья Google Scholar
A. Consiglio, A. Borsoi, G. Pereira, A. Lucion, Влияние окситоцина, вводимого в центральное ядро миндалины и ядро ложа stria terminalis, на агрессивное поведение матери у крыс. Physiol. Behav. 85 (3), 354–362 (2005)
CAS PubMed Статья Google Scholar
С. Кенни, Л. Райт, А. Грин, Р. Масудх, Т. Перро, Выражение материнского поведения и активация ядра ложа терминальной полоски во время воздействия угрозы хищников: модулирующие эффекты транспортного стресса .Physiol. Behav. 17 (123), 148–155 (2014)
Статья CAS Google Scholar
Мангуриан Л.П., Р.Дж. Уолш, Б. Познер, Повышение собственного поглощения пролактина сосудистым сплетением. Эндокринология 131 , 698–702 (1992)
CAS PubMed Google Scholar
R.A. Августин, И. Кокай, З.Б. Эндрюс, С. Ладиман, Д.Р. Граттан, Количественное определение мРНК рецептора пролактина в головном мозге матери крысы во время беременности и кормления грудью. J. Mol. Эндокринол. 31 (1), 221–232 (2003)
CAS PubMed Статья Google Scholar
Х. Табата, М. Кобаяши, Дж. Х. Икеда, Н. Накао, Т. Сайто, М. Танака, Характеристика нескольких первых экзонов в гене рецептора мышиного пролактина и влияние пролактина на их экспрессию в сосудистом сплетении.J. Mol. Эндокринол. 48 (2), 169–176 (2012)
CAS PubMed Статья Google Scholar
X.J. Пи, Д. Граттан, Дифференциальная экспрессия двух форм мРНК рецептора пролактина в микродиссектированных ядрах гипоталамуса крысы. Brain Res. Мол. Brain Res. 59 (1), 1–12 (1998)
CAS PubMed Статья Google Scholar
Р. Абс, Э. Ван Влеймен, П. Паризель, К. Ван Аккер, М. Мартин, Дж. Мартин, Врожденная гипоплазия мозжечка и гипогонадотропный гипогонадизм. J. Neurol. Sci. 98 (2–3), 259–265 (1990)
CAS PubMed Статья Google Scholar
Г. Ленг, Р. Дайболл, С. Лакман, Механизмы секреции вазопрессина. Horm. Res. 37 (1–2), 33–38 (1992)
CAS PubMed Статья Google Scholar
Г. Кеннеди, Гипоталамический контроль потребления пищи у крыс. Proc. R. Soc. Лондон. B. Biol. Sci. 137 (889), 535–549 (1950)
CAS PubMed Статья Google Scholar
К.М., Брукс, Э., Ламберт, П., Бард, Экспериментальное производство ожирения у обезьяны (Macaca mulatta). Кормили. Proc. 1 , 11 (1942)
Д. Сове, Б. Вудсайд, Влияние центрального введения пролактина на потребление пищи девственными самками крыс зависит от дозы, происходит в отсутствие гормонов яичников и латентность до начала заболевания зависит от режима кормления.Brain Res. 729 (1), 75–81 (1996)
PubMed Статья Google Scholar
Д. Сове, Б. Вудсайд, Нейроанатомическая специфичность пролактин-индуцированной гиперфагии у девственных самок крыс. Brain Res. 868 (2), 306–314 (2000)
CAS PubMed Статья Google Scholar
S. Mejía, L.M. Torner, M.C. Езёрски, К. Гонсалес, М.А. Моралес, Г. Мартинес де ла Эскалера, Ч .: Клапп, пролактин и пролактин 16K стимулируют высвобождение вазопрессина путем прямого воздействия на гипоталамо-нейрогипофизарную систему. Эндокринная 20 (1–2), 155–162 (2003)
PubMed Статья Google Scholar
С. Мехиа, М. Моралес, М. Зетина, Г. Мартинес де ла Эскалера, Ч .: Клапп, Иммунореактивные формы пролактина совместно с вазопрессином локализуются в нейронах паравентрикулярных и супраоптических ядер гипоталамуса.Нейроэндокринология 66 (3), 151–159 (1997)
PubMed Статья Google Scholar
К. Педерсен, Дж. Ашер, Ю. Монро, А.Дж. Прейндж, окситоцин вызывает материнское поведение у девственных самок крыс. Наука 216 (4546), 648–650 (1982)
CAS PubMed Статья Google Scholar
J.M. Zimmermann-Peruzatto, V.M. Лаццари, А.К. де Моура, С. Алмейда, М. Джовенарди, Изучение роли вазопрессина в модуляции родительского и сексуального поведения. Фронт. Психиатрия 6 , 1–8 (2015)
Статья Google Scholar
H.H. Van Tol, E.L. Болверк, Б. Лю, Дж. П. Бурбах, Экспрессия генов окситоцина и вазопрессина в гипоталамо-нейрогипофизарной системе крыс во время эстрального цикла, беременности и кормления грудью. Эндокринология 122 (3), 945–951 (1988)
PubMed Статья Google Scholar
Н. Доннер, И. Нойман, Влияние хронического внутримозгового пролактина на окситоцинергическую и вазопрессинергическую систему у девственных овариэктомированных крыс. Нейроэндокринология 90 (3), 315–322 (2009)
CAS PubMed Статья Google Scholar
Р., Августин, Г., Бауэр, А., Сеймур, Д., Граттан, С., Браун, Репродуктивная регуляция экспрессии генов в супраоптических и паравентрикулярных ядрах гипоталамуса.J. Neuroendocrinol. (4), (2016). DOI: 10.1111 / jne.12350
К. Судзуки, Н. Коидзуми, Х. Хиросе, Р. Хокао, Н. Такемура, С. Мотоёси, Изменения в плазме концентрации аргинина вазопрессина во время лактации у крыс. Комп. Med. 50 (3), 277–280 (2000)
CAS PubMed Google Scholar
N. Donner, R. Bredewold, R. Maloumby, I.D. Neumann, Хронический внутримозговый пролактин ослабляет нервные цепи стресса у девственных крыс.Евро. J. Neurosci. 25 (6), 1804–1814 (2007)
PubMed Статья Google Scholar
Л. Крулич, Э. Хефко, П. Илнер, К. Б. Рид, Влияние острого стресса на секрецию ЛГ, ФСГ, пролактина и ГР у нормальных самцов крыс, с комментариями по их статистической оценке. Нейроэндокринология 16 , 293–311 (1974)
CAS PubMed Статья Google Scholar
Б. Бодози, Ф. Обал мл., Дж. Гарди, Дж. Комлоди, Дж. Фанг, Дж. М. Крюгер, Эфирный стрессор увеличивает быстрый сон у крыс: возможная роль пролактина. Являюсь. J. Physiol. 279 (5), 1590–1589 (2000)
Google Scholar
Р.Р. Гала, Физиология и механизмы стресс-индуцированных изменений секреции пролактина у крыс. Life Sci. 46 , 1407–1420 (1990)
CAS PubMed Статья Google Scholar
Ф. Обал мл., М. Опп, А. Б. Cady, L. Johannsen, J.M. Krueger, пролактин, вазоактивный кишечный пептид и пептид гистидинметионин вызывают избирательное увеличение фазы быстрого сна у кроликов. Brain Res. 490 , 292–300 (1989)
CAS PubMed Статья Google Scholar
P. Meerlo, A. Easton, B.M. Бергманн, Ф.В. Турек, Сдерживание увеличивает пролактин и быстрый сон у мышей C57BL / 6J, но не у мышей BALB / cJ.Являюсь. J. Physiol. 281 (3), 846–854 (2001)
Google Scholar
Р. Б. Мачадо, М. Р. Роча, Д. Сучеки, Пролактин мозга участвует в индуцированном стрессом восстановлении быстрого сна. Horm. Behav. 89 , 38–47 (2016)
PubMed Статья CAS Google Scholar
Р.М. Frieboes, H. Murck, G.K. Сталла, И. Антониевич, А. Штайгер, Улучшенный медленный сон у пациентов с пролактиномой.J. Clin. Эндокринол. Метаб. 83 , 2706–2710 (1998)
CAS PubMed Статья Google Scholar
К. Коике, А. Мияке, Т. Аоно, Т. Сакумото, М. Омичи, М. Ямагути, О. Танидзава, Влияние пролактина на секрецию гипоталамического ГнРГ и гонадотропинов гипофиза. Horm. Res. 35 (1), 5–12 (1991)
CAS PubMed Статья Google Scholar
Р. Араужо-Лопес, Дж. Крэмптон, Н. Акино, Р. Миранда, И. Кокай, А. Рейс, К. Р. Фрэнси, Д. Граттан, Р. Szawka, Пролактин регулирует нейроны кисспептина в дугообразном ядре, подавляя секрецию ЛГ у самок крыс. Эндокринология 155 (3), 1010–1020 (2014)
PubMed Статья CAS Google Scholar
Дж. Кларксон, X. д’Англемон де Тассиньи, В. Колледж, А. Карати, А. Гербисон, Распределение нейронов кисспептина в мозге взрослых самок мышей.J. Neuroendocrinol. 21 (8), 673–682 (2009)
CAS PubMed Статья Google Scholar
I.C. Кокай, С. Петерсен, Д. Граттан, Идентификация пролактин-чувствительных нейронов ГАМК и кисспептина в областях гипоталамуса крыс, участвующих в контроле фертильности. Эндокринология 152 (2), 526–535 (2011)
CAS PubMed Статья Google Scholar
I. Cohen-Becker, M. Selmanoff, P. Wise, Гиперпролактинемия изменяет частоту и амплитуду пульсирующей секреции лютеинизирующего гормона у овариэктомированных крыс. Нейроэндокринология 42 (4), 328–333 (1986)
CAS PubMed Статья Google Scholar
A. Lerant, M.E. Freeman, яичниковые стероиды по-разному регулируют экспрессию PRL-R в популяциях нейроэндокринных дофаминергических нейронов: конфокальное микроскопическое исследование с двойной меткой.Brain Res. 802 (1–2), 141–154 (1998)
CAS PubMed Статья Google Scholar
Г. Андерсон, Д. Граттан, В. ван ден Анкер, Р. Бриджес, Репродуктивный опыт увеличивает чувствительность к пролактину в медиальной преоптической области и дугообразном ядре самок крыс. Эндокринология 147 , 4688–4694 (2006)
CAS PubMed Статья Google Scholar
L. Torner, R. Maloumby, G. Nava, J. Aranda, C. Clapp, I.D. Neumann, Высвобождение in vivo и активация гена пролактина мозга в ответ на физиологические стимулы. Евро. J. Neurosci. 19 (6), 1601–1608 (2004)
PubMed Статья Google Scholar
J.A. МакГенри, Д. Рубинов, Г.Д. Стубер, Материнские ответные нейроны в ядре ложа терминальной полоски и медиальной преоптической области: предполагаемые схемы для регулирования тревоги и вознаграждения.Фронт. Нейроэндокринол. 38 , 65–72 (2015)
PubMed PubMed Central Статья Google Scholar
Р.С. Бриджес, М. Нуман, П.М. Ронсхайм, П. Mann, C.E. Lupini, Центральные инфузии пролактина стимулируют материнское поведение у нерожавших самок крыс, получавших стероиды. Proc. Natl. Акад. Sci. США 87 (20), 8003–8007 (1990)
CAS PubMed PubMed Central Статья Google Scholar
F.K. Стефан, И. Цукер, Циркадные ритмы питьевого поведения и двигательной активности крыс устраняются поражениями гипоталамуса. Proc. Natl. Акад. Sci. США 69 (6), 1583–1586 (1972)
CAS PubMed PubMed Central Статья Google Scholar
C. Bethea, J. Neill, Поражения супрахиазматических ядер устраняют у крыс выбросы пролактина, стимулированные цервикально. Эндокринология 107 (1), 1–5 (1980)
CAS PubMed Статья Google Scholar
К. Браун-Грант, Г. Райсман, Нарушения репродуктивной функции, связанные с разрушением супрахиазматических ядер у самок крыс. Proc. R. Soc. Лондон. B Biol. Sci. 198 (1132), 279–296 (1977)
CAS PubMed Статья Google Scholar
J.T. Пан, Р.Р.Гала, Области центральной нервной системы, вовлеченные в вызванный эстрогеном всплеск пролактина во второй половине дня. II. Имплантационные исследования. Эндокринология 117 (1), 388–395 (1985)
CAS PubMed Статья Google Scholar
Дж. Пан, Р. Гала, Области центральной нервной системы, вовлеченные в вызванный эстрогеном выброс пролактина во второй половине дня. I. Исследования повреждений. Эндокринология 117 (1), 382–387 (1985)
CAS PubMed Статья Google Scholar
И. Кларк, Дж. Камминс, Д. де Крецер, Функция гипофиза у овец после отключения от прямых гипоталамических влияний. Нейроэндокринология 36 (5), 376–384 (1983)
CAS PubMed Статья Google Scholar
J.D. Johnston, D.J. Скене, Регуляция нейроэндокринной физиологии и ритмов млекопитающих с помощью мелатонина. J. Endocrinol. 226 (2), T187 – T198 (2015)
CAS PubMed Статья Google Scholar
С. Шир, В. Кроули, С. Гросвенор, Влияние неонатальной недостаточности пролактина на препубертатный тубероинфундибулер и туберогипофизарную дофаминергическую нейронную активность. Эндокринология 119 (3), 1217–1221 (1986)
CAS PubMed Статья Google Scholar
C. Grosvenor, N. Whitworth, Накопление пролактина в материнском молоке и его передача в кровообращение новорожденных крыс: обзор. Эндокринол. Exp. 17 , 271–282 (1983)
CAS PubMed Google Scholar
М. Ройстер, П. Дрисколл, П. Келли, М. Фримарк, Рецептор пролактина у плода крысы: клеточная локализация мессенджера рибонуклеиновой кислоты, иммунореактивного белка, лиганд-связывающей активности и индукции экспрессии в поздняя беременность.Эндокринология 136 (9), 3892–3900 (1995)
CAS PubMed Статья Google Scholar
А.И. Melo, M. Perez-Ledezma, C. Clapp, E. Arnold, J.C. Rivera, A.S. Флеминг, Влияние дефицита пролактина в раннем постнатальном периоде на развитие материнского поведения у самок крыс: материнское молоко имеет значение. Horm. Behav. 56 (3), 281–291 (2009)
CAS PubMed Статья Google Scholar
E. Vergara-Castañeda, D.R. Граттан, Х. Пасантес-Моралес, М. Перес-Домингес, Э.А. Кабрера-Рейес, Т. Моралес, М. Цербон, Пролактин опосредует нейрозащиту против эксайтотоксичности в первичных культурах клеток нейронов гиппокампа через свой рецептор. Brain Res. 1636 , 192–199 (2016)
Артикул CAS Google Scholar
N.V. Emanuele, L. Metcalfe, L. Wallock, J. Tentler, T.C. Хаген, К. Бир, Д. Мартинсон, П.W. Gout, L. Kirsteins, A.M. Лоуренс, Внегипонталамический пролактин головного мозга: характеристика и доказательства независимости от пролактина гипофиза. Brain Res. 421 (1–2), 255–262 (1987)
CAS PubMed Статья Google Scholar
Х. Эйхенбаум, Гиппокамп: когнитивные процессы и нейронные репрезентации, лежащие в основе декларативной памяти. Нейрон 44 (1), 109–120 (2004)
CAS PubMed Статья Google Scholar
Г. Лав, Н. Торри, И. Макнамара, М. Морган, М. Бэнкс, Н. Хестер, Э. Р. Гласпер, A.C. Devries, C.H. Кинсли, К. Ламберт, Материнский опыт вызывает у крыс долговременные изменения в поведении. Behav. Neurosci. 119 (4), 1084–1096 (2005)
PubMed Статья Google Scholar
C.H. Кинсли, Л. Мадония, Г. Гиффорд, К. Турески, Г. Гриффин, К. Лоури, Дж. Уильямс, Дж. Коллинз, Х. Маклери, К.Г. Ламберт, Материнство улучшает обучение и память. Nature 402 (6758), 137–138 (1999)
CAS PubMed Статья Google Scholar
J.L. Pawluski, B.L. Вандербил, К. Раган, Л.А. Галеа, Первый репродуктивный опыт постоянно влияет на пространственную привязку и рабочую память у матери, и эти эффекты возникают не только из-за беременности или «материнства». Behav. Brain Res. 175 (1), 157–165 (2006)
PubMed Статья Google Scholar
J.L. Pawluski, S.K. Уокер, Л.А. Галеа, Репродуктивный опыт по-разному влияет на пространственную привязку и производительность рабочей памяти у матери. Horm. Behav. 49 (2), 143–149 (2006)
PubMed Статья Google Scholar
Дж. Генри, Б. Шервин, Гормоны и когнитивные функции на поздних сроках беременности и в послеродовом периоде: продольное исследование. Behav. Neurosci. 126 (1), 73–85 (2012)
CAS PubMed Статья Google Scholar
Т. Кастаньо, П. Морейра, К. Португал-Нуньес, А. Новаис, П. Коста, Дж. Палха, Н. Соуза, Н. К. Сантос, Роль пола и связанных с полом гормонов в познании, настроении и благополучии. бытие у пожилых мужчин и женщин. Биол. Psychol. 103 , 158–166 (2014)
PubMed Статья Google Scholar
T.L. Уокер, Я. Вукович, М. Кудийс, Д. Блэкмор, E.W. Mackay, A.M. Сайкс, Р.В. В целом, A.S. Хэмлин, П.Ф. Бартлетт, Пролактин стимулирует клетки-предшественники в гиппокампе взрослых мышей.PLoS One 7 (9), 1–11 (2012)
Артикул CAS Google Scholar
Дж. Рейес-Мендоза, Т. Моралес, Последующая обработка пролактином защищает нейроны СА1 гиппокампа овариэктомированной самки крысы от нейродегенерации, вызванной каиновой кислотой. Неврология 328 , 58–68 (2016)
CAS PubMed Статья Google Scholar
J.L. Pawluski, L.A. Galea, Репродуктивный опыт изменяет нейрогенез гиппокампа в послеродовом периоде у матери. Неврология 149 (1), 53–67 (2007)
CAS PubMed Статья Google Scholar
Г. Мак, С. Вайс, Отцовское распознавание взрослых потомков, опосредованное вновь созданными нейронами ЦНС. Nat. Neurosci. 13 (6), 753–758 (2010)
CAS PubMed Статья Google Scholar
Г.В. Pardo, J.F. Goularte, A.L. Hoefel, A.L. de Castro, L.C. Кухарски, А. да Роса Араужо, А.Б .: Люсьон, Влияние ограничения сна во время беременности на мать и плод у крыс. Physiol. Behav. 1 (155), 66–76 (2016)
Статья CAS Google Scholar
Г. Мак, Э. Энвере, К. Грегг, Т. Пакарайнен, М. Поутанен, И. Хухтаниеми, С. Вайс, Нейрогенез, стимулированный мужскими феромонами в мозге взрослой женщины: возможная роль в брачном поведении .Nat. Neurosci. 10 (8), 1003–1011 (2007)
CAS PubMed Статья Google Scholar
Э. Гулд, Б.С. МакИвен, П. Танапат, Л.А. Галеа, Э. Фукс, Нейрогенез в зубчатой извилине взрослой бурозубки регулируется психосоциальным стрессом и активацией рецепторов NMDA. J. Neurosci. 17 (7), 2492–2498 (1997)
CAS PubMed Google Scholar
Р.С. Думан, Дж. Мальберг, С. Накагава, Регулирование нейрогенеза взрослых с помощью психотропных препаратов и стресса. J. Pharmacol. Exp. Ther. 299 (2), 401–407 (2001)
CAS PubMed Google Scholar
E.A. Нимчинский, Б. Сабатини, К. Свобода, Строение и функция дендритных шипов. Анну. Rev. Physiol. 64 , 313–353 (2001)
Артикул Google Scholar
C.H. Кинсли, Р. Трейнер, Г. Стафиссо-Сандос, П. Куадрос, Л.К. Маркус, К. Херон, Э.А. Мейер, Н. Хестер, М. Морган, Ф.Дж. Козуб, К.Г. Ламберт, Материнство и гормоны беременности изменяют концентрацию нейрональных дендритных шипов гиппокампа. Horm. Behav. 49 (2), 131–142 (2006)
CAS PubMed Статья Google Scholar
J.L. Pawluski, L.A. Galea, На морфологию гиппокампа по-разному влияет репродуктивный опыт матери.J. Neurobiol. 66 (1), 71–81 (2006)
PubMed Статья Google Scholar
J. Brusco, R. Wittmann, M. de Azevedo, A. Lucion, C. Franci, M. Giovenardi, A.A. Rasia-Filho, Гормональные профили плазмы, плотность и морфология дендритных шипов в лучевом слое CA1 гиппокампа, полученные с помощью световой микроскопии, у девственных и послеродовых самок крыс. Neurosci. Lett. 438 (3), 346–350 (2008)
CAS PubMed Статья Google Scholar
Р. Аббуд, Г. Хоффман, М. Смит, Рефрактерность коры к стимуляции N-метил-D, L-аспарагиновой кислоты (NMDA) у кормящих крыс: восстановление после удаления детенышей и блокада рецепторов прогестерона. Brain Res. 604 (1–2), 16–23 (1993)
CAS PubMed Статья Google Scholar
C.A. Стендли, связывание рецептора N-метил-D-аспартата изменено, и судорожный потенциал снижен у беременных крыс. Brain Res. 844 (1–2), 10–19 (1999)
CAS PubMed Статья Google Scholar
П. Анборг, К. Годин, М. Пампилло, Г. Дами, Л. Дейл, С. Креган, Р. Труант, С. С. Фергюсон, Ингибирование передачи сигналов метаботропного рецептора глутамата с помощью связывающего гентингтин белка оптинеурина. . J. Biol. Chem. 280 (41), 34840–34848 (2005)
CAS PubMed Статья Google Scholar
М.Р. Хайнд, Х.Л. Скотт, П.Р. Додд, Глутамат-опосредованная эксайтотоксичность и нейродегенерация при болезни Альцгеймера. Neurochem. Int. 45 (5), 583–595 (2004)
CAS PubMed Статья Google Scholar
М. Доретто, М. Оливейра-э-Силва, Д. Феррейра-Алвес, С. Пирес, Н. Гарсиа-Кайраско, А. Рейс, Влияние лактации на проявление аудиогенных приступов: связь с профили пролактина плазмы.Epilepsy Res. 54 , 109–121 (2003)
CAS PubMed Статья Google Scholar
P. Berzaghi Mda, D. Amado, E. Cavalheiro, Беременность снижает частоту спонтанных рецидивирующих припадков у крыс с поражением гиппокампа каиновой кислотой. Epilepsy Res. 1 (2), 142–144 (1987)
PubMed Статья Google Scholar
Т. Моралес, М. Лоренсон, А.М. Уокер, Э. Рамос. Как пролактин (PRL), так и молекулярный имитатор фосфорилированного PRL, S179D-PRL, защищают гиппокамп самок крыс от эксайтотоксичности. Неврология 258 , 211–217 (2014)
CAS PubMed Статья Google Scholar
Н.А. Риверо-Сегура, Э. Флорес-Сото, С. Гарсия де ла Кадена, И. Коронадо-Марес, Х.С. Гомес-Верьян, Д.Г. Феррейра, Э.А. Кабрера-Рейес, Л.V. Lopes, L. Massieu, M .: Cerbón, Пролактин-индуцированная нейрозащита от эксайтотоксичности глутамата опосредуется снижением перегрузки [Ca2 +] i и активации NF-κB. PLoS One 12 (5), e0176910 (2017)
PubMed PubMed Central Статья Google Scholar
A.M. Томсон, Неокортикальный слой 6, обзор. Фронт. Нейроанат. 4 , 13 (2010)
PubMed PubMed Central Google Scholar
И.Ю. Чжоу, Р.В. Чан, Л.С. Хо, Э. Ву, Продольные метаболические изменения в гиппокампе и таламусе материнского мозга, выявленные с помощью протонной магнитно-резонансной спектроскопии. Neurosci. Lett. 553 , 170–175 (2013)
CAS PubMed Статья Google Scholar
J. Broida, S.D. Michael, B. Svare, Уровни пролактина плазмина не связаны с инициированием, поддержанием и снижением послеродовой агрессии у мышей.Behav. Neural. Биол. 32 (1), 121–125 (1981)
CAS PubMed Статья Google Scholar
М. Гарланд, Б. Сваре, Стимуляция сосания модулирует поддержание послеродовой агрессии у мышей. Physiol. Behav. 44 (3), 301–305 (1988)
CAS PubMed Статья Google Scholar
O.J. Bosch, I.D. Нейман, вазопрессин, высвобождаемый в центральной миндалине, способствует материнской агрессии.Евро. J. Neurosci. 31 (5), 883–891 (2010)
PubMed Статья Google Scholar
Дэвис М. Роль миндалины при страхе и тревоге. Анну. Rev. Neurosci. 15 , 353–375 (1992)
CAS PubMed Статья Google Scholar
Дж. Аллен, К. Аллен, Роль миндалоидных комплексов в вызванном стрессом высвобождении АКТГ у крыс.Нейроэндокринология 15 (3–4), 220–230 (1974)
CAS PubMed Google Scholar
Н. Доннер, Р. Бредеволд, Р. Малумби, И. Нойман, Хронический внутримозговый пролактин ослабляет нервные цепи стресса у девственных крыс. Евро. J. Neurosci. 25 (6), 1804–1814 (2007)
PubMed Статья Google Scholar
Л. Торнер, Н.Toschi, A. Pohlinger, R. Landgraf, I.D. Нойман, Анксиолитические и антистрессовые эффекты мозгового пролактина: повышение эффективности антисмыслового воздействия на рецептор пролактина путем молекулярного моделирования. J. Neurosci. 21 (9), 3207–3214 (2001)
CAS PubMed Google Scholar
О. Пиказо, А. Фернандес-Гуасти, Изменения в экспериментальной тревожности во время беременности и кормления грудью. Physiol. Behav. 54 (2), 295–299 (1993)
CAS PubMed Статья Google Scholar
D. Toufexis, S. Tesolin, N. Huang, C. Walker, Измененная чувствительность гипофиза к кортикотропин-рилизинг-фактору и аргинин-вазопрессину участвует в стрессовой гипореактивности лактации у крыс. J. Neuroendocrinol. 11 (10), 757–764 (1999)
CAS PubMed Статья Google Scholar
L.D. Сэнфорд, П. Нассар, Р.Дж. Росс, Дж. Шулькин, А. Моррисон, Микроинъекции пролактина в центральное ядро миндалины приводят к снижению NREM-сна.Sleep Res. Интернет 1 , 109–113 (1998)
CAS PubMed Google Scholar
X. Pi, J.L. Voogt, D.R. Граттан, Обнаружение мРНК рецептора пролактина в полосатом теле и черном веществе крысы. J. Neurosci. Res. 67 (4), 551–558 (2002)
CAS PubMed Статья Google Scholar
B.D. Пальма, Д.К. Гиполид, С.Туфик, Влияние на секрецию пролактина и связывание с дофаминергическими рецепторами у мышей, лишенных сна, склонных к волчанке. Braz. J. Med. Биол. Res. 42 (3), 299–304 (2009)
CAS PubMed Статья Google Scholar
П. Малвен, Ингибирование высвобождения пролактина у овец в сознании после стимуляции прилежащего ядра и хвостатого ядра. Нейроэндокринология 28 (3), 160–168 (1979)
CAS PubMed Статья Google Scholar
R.S.E. Браун, А.Э. Гербисон, Д. Граттан, Дифференциальные изменения в ответах популяций нейронов гипоталамуса и ствола мозга на пролактин во время лактации у мышей. Биол. Репродукция. 84 , 826–836 (2011)
CAS PubMed Статья Google Scholar
Л. Калигарис, С. Талейсник, Участие нейронов, содержащих 5-гидрокситриптамин, в механизме высвобождения пролактина, индуцированного эстрогеном. Дж.Нейроэндокринол. 62 (1), 25–33 (1974)
CAS Google Scholar
Г.А. Ян, Р.П. Деис, Влияние антагонистов серотонина на секрецию пролактина и прогестерона у крыс: данные о том, что стимулирующее и ингибирующее действие серотонина на высвобождение пролактина может опосредоваться различными рецепторами. J. Endocrinol. 117 (3), 415–422 (1988)
CAS PubMed Статья Google Scholar
А. Мистри, Дж. Л. Вугт, Роль серотонина в ночных и суточных выбросах пролактина у беременных крыс. Эндокринология 125 (6), 2875–2880 (1989)
CAS PubMed Статья Google Scholar
Л. Паут-Пагано, Р. Роки, Дж. Валаткс, К. Китахама, М. Жуве, Анатомическое распределение пролактин-подобной иммунореактивности в головном мозге крыс. Нейроэндокринология 58 (6), 682–695 (1993)
CAS PubMed Статья Google Scholar
М. Ангоа-Перес, Д. Кун, Нейрональный серотонин в регуляции материнского поведения у грызунов. Нейротрансмиттер 2 , 615–625 (2015)
Google Scholar
M.C. Попеско, А. Фростхольм, К. Рейняк, А. Роттер, Цифровой анализ транскриптома в стареющем мозжечке. Аня. N.Y. Acad. Sci. 1019 , 58–63 (2004)
CAS PubMed Статья Google Scholar
А.Л. Барофски, Дж. Тейлор, В.Дж. Massari, Выступы спинного шва и гипоталамуса обеспечивают стимулирующий серотонинергический сигнал для высвобождения пролактина, вызванного грудным вскармливанием. Эндокринология 113 (5), 1894–1903 (1983)
CAS PubMed Статья Google Scholar
П. Эрикссон, Э. Перфильева, Т. Бьорк-Эрикссон, А. Альборн, К. Нордборг, Д. Петерсон, Ф. Х. Гейдж, Нейрогенез в гиппокампе взрослого человека. Nat. Med. 4 (11), 1313–1317 (1998)
CAS PubMed Статья Google Scholar
Пролактин — Функции — Регулирование — Секреция
Пролактин — один из гормонов, вырабатываемых передней долей гипофиза. Он выполняет множество функций, но особенно важен для развития груди и производства грудного молока у женщин.
В этой статье мы обсудим пролактин, то, как регулируется его секреция, и соответствующие клинические условия.
Производство пролактина
Пролактин (PRL) продуцируется лактотрофными клетками передней доли гипофиза. Гипоталамус не производит стимулирующих гормонов.
Функции пролактина
Пролактин обладает хорошо известными физиологическими функциями только у женщин. Постепенное увеличение ПРЛ во время беременности, наряду с высокими уровнями эстрогена и прогестерона, приводит к полноценному развитию груди.
Основные действия пролактина перечислены ниже:
- Запуск производства молока альвеолярными клетками (лактогенный)
- Поддержание производства молока после того, как оно было установлено (галактоопоэтическое)
- Пролиферация альвеолярных и протоковых клеток (маммогенная)
Пролактин в первую очередь оказывает лактогенное и галактоопоэтическое действие.
Рис. 1. Схема, показывающая структуру кормящей груди [/ caption]Регулирование секреции пролактина
Высвобождение пролактина ингибируется дофамином (DA) , известным в данном контексте как фактор ингибирования ПРЛ (PIF) .
Сосание — самый мощный стимул для выброса ПРЛ. Стимуляция соска посылает сигналы через афферентных нервных путей вверх по спинному мозгу. Это подавляет дофаминергические нейроны в гипоталамусе.Таким образом, происходит ингибирование тормозящего нейромедиатора. Это называется растормаживанием.
И наоборот, тиреотропин-рилизинг-гормон (TRH) , также вырабатываемый гипоталамусом, стимулирует высвобождение пролактина. Кроме того, эстрогены повышают чувствительность лактотрофов к TRH, а также снижают их чувствительность к ингибированию дофамина.
Рис. 2. Диаграмма, показывающая регуляцию секреции пролактина [/ caption]Примечание: ось пролактина технически не имеет системы отрицательной обратной связи.Без ингибирования дофамином пролактин будет секретироваться бесконечно.
[старт-клиника]
Клиническая значимость — грудное вскармливание
Лактация обычно подавляет нормальный цикл яичников. Уменьшает высвобождение GnRH нейронами гипоталамуса. Это вызывает снижение высвобождения ФСГ и ЛГ, которые отвечают за нормальную функцию яичников.
Кормление грудью не прекращает на неопределенный срок яичниковые циклы, и в конечном итоге они возобновятся независимо от того, кормят ребенка грудью или нет.Если мать не кормит ребенка грудью, овуляторные циклы обычно возобновляются через 8-10 недель после родов.
Клиническая значимость — Гиперпролактинемия
Гиперпролактинемия — это состояние повышенного уровня пролактина на . Это может вызвать:
- Нарушения менструального цикла — аменорея или олигоменорея
- Галакторея (подтекание молока через соски)
- гипогонадизм, вызывающий бесплодие и эректильную дисфункцию, а также остеопороз
Причины можно запомнить как 5 Ps:
- P гизиологический — кормление грудью, стресс, острые подъемы после полового акта
- P регнантство
- P хармакологический e.грамм. антагонисты дофамина, нейролептики (из-за эффектов антагонистов дофамина)
- P ролактинома (пролактинпродуцирующая опухоль гипофиза)
- P Синдром кистозных яичников (СПКЯ). Механизм точно не известен, но предполагается, что он связан с повышенным уровнем эстрогена, оказывающим стимулирующее действие на выработку пролактина
Диагноз основывается на обнаружении первопричины, и лечение будет подбираться соответствующим образом.
[конец клинической]
Базальная функция пролактина костистых рыб как ключевого регулятора захвата ионов идентифицирована с нокаут-моделями рыбок данио
Riddle, O., Бейтс, Р. В. и Дикшорн, С. В. Подготовка, идентификация и анализ пролактина — гормона передней доли гипофиза. Am J Physiol 105, 191–216 (1933).
CAS Google Scholar
Ormandy, C.J. et al. Нулевая мутация гена рецептора пролактина вызывает множественные репродуктивные дефекты у мышей. Гены и развитие 11, 167–178 (1997).
CAS Google Scholar
Веннбо, Х.и другие. Активация рецептора пролактина, но не рецептора гормона роста, важна для индукции опухолей молочной железы у трансгенных мышей. Журнал клинических исследований 100, 2744–2751 (1997).
CAS PubMed PubMed Central Google Scholar
Бен-Джонатан, Н., ЛаПензее, К. Р. и ЛаПенси, Э. У. Что мы можем узнать от грызунов о пролактине у людей? Endocr Rev 29, 1–41 (2008).
CAS PubMed Google Scholar
Всадник, Н.Д. и Грегерсон, К. А. Действие пролактина. J Mol Endocrinol 52, R95–106 (2014).
CAS PubMed Google Scholar
Уиттингтон, К. М. и Уилсон, А. Б. Роль пролактина в воспроизводстве рыб. Gen Comp Endocrinol 191, 123–136 (2013).
CAS PubMed Google Scholar
Манзон, Л. А. Роль пролактина в осморегуляции рыб: обзор.Gen Comp Endocrinol 125, 291–310 (2002).
CAS PubMed Google Scholar
Пикфорд, Г. Э. и Филлипс, Дж. Г. Пролактин, фактор, способствующий выживанию гипофизэктомированных киллеров в пресной воде. Science 130, 454–455 (1959).
ADS CAS PubMed Google Scholar
Мадсен, С. С., Дженсен, М. К., Нор, Дж. И Кристиансен, К. Экспрессия Na + -K + -АТФазы в кумже, Salmo trutta: In vivo Модуляция гормонами и морской водой.Am J Physiol-Reg I 269, R1339 – R1345 (1995).
CAS Google Scholar
Сил, А. П. и др. Влияние солености и пролактина на уровни транскриптов генов переносчиков ионов, ионных насосов и рецепторов пролактина в кишечнике тилапии Мозамбика. Gen Comp Endocrinol 206, 146–154 (2014).
CAS PubMed Google Scholar
Сакамото, Т. и Маккормик, С.D. Пролактин и гормон роста в осморегуляции рыб. Gen Comp Endocrinol 147, 24–30 (2006)
CAS PubMed Google Scholar
Хван, П. П. и Чжоу, М. Ю. Рыбки данио как модель на животных для изучения ионного гомеостаза. Pflug Arch Eur J Phy 465, 1233–1247 (2013).
CAS Google Scholar
Huang, X. G. et al. Открытие нового пролактина у позвоночных, не являющихся млекопитающими: эволюционные перспективы и его участие в развитии сетчатки костистых мышц.Plos One 4, e6163 (2009).
ADS PubMed PubMed Central Google Scholar
Бревес, Дж. П., Серизье, С. Б., Гоффин, В., Маккормик, С. Д. и Карлстром, Р. О. Пролактин регулирует транскрипцию гена захвата ионов Na + / Cl- котранспортера (ncc) в жабрах рыбок данио. Mol Cell Endocrinol 369, 98–106 (2013).
CAS PubMed PubMed Central Google Scholar
Чжу, Ю., Сонг, Д., Тран, Н. Т. и Нгуен, Н. Эффекты нокдауна членов семейства гормонов роста в развитии рыбок данио. Gen Comp Endocrinol 150, 395-404 (2007).
CAS PubMed Google Scholar
Chasiotis, H. & Kelly, S.P. Иммунолокализация окклюдина и экспрессия белка у золотых рыбок. J Exp Biol 211, 1524–1534 (2008).
CAS PubMed Google Scholar
Cerda, J.& Финн, Р. Н. Писцин Аквапорины: Обзор последних достижений. Журнал J Exp Zool, часть A 313A, 623–650 (2010).
CAS Google Scholar
Hsiao, C.D. et al. Положительная регуляторная петля между foxi3a и foxi3b важна для спецификации и дифференцировки эпидермальных ионоцитов рыбок данио. Plos One 2, e302 (2007).
ADS PubMed PubMed Central Google Scholar
Чанг, W.Дж. И Хванг, П. П. Развитие эпидермиса рыбок данио. Исследование врожденных дефектов, часть C-Embryo Today 93, 205–214 (2011).
CAS Google Scholar
Кумай, Ю., Квонг, Р. В. и Перри, С. Ф. Роль цАМФ-опосредованной внутриклеточной передачи сигналов в регулировании захвата Na + у личинок рыбок данио. Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология 306, R51–60 (2014).
CAS PubMed Google Scholar
Чанг, W.J. et al. Компенсаторная регуляция поглощения Na + обменником Na + / H + и котранспортером Na + -Cl- у рыбок данио (Danio rerio). Фронт Зоол 10 (2013).
PubMed PubMed Central Google Scholar
Wang, YF, Tseng, YC, Yan, JJ, Hiroi, J. & Hwang, PP Роль SLC12A10.2, белка, подобного котранспортеру Na-Cl, в механизме захвата Cl у рыбок данио (Danio rerio) . Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология 296, R1650–1660 (2009).
CAS PubMed Google Scholar
Ляо, Б.К., Чен, Р.Д. и Хванг, П.П. Регулирование экспрессии подтипов альфа1-субъединицы Na + -K + -АТФазы в жаберных ионоцитах рыбок данио. Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология 296, R1897–1906 (2009).
CAS PubMed Google Scholar
Mobasheri, A. et al. Разнообразие изоферментов Na +, K + -АТФазы; Сравнительная биохимия и физиологические последствия новых функциональных взаимодействий.Bioscience Rep 20, 51–91 (2000).
CAS Google Scholar
Квонг, Р. В. и Перри, С. Ф. Белок плотных контактов клаудин-b регулирует проницаемость эпителия и обработку натрия у личинок рыбок данио, Danio rerio. Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология 304, R504–513 (2013).
CAS PubMed PubMed Central Google Scholar
Деспа, С.И Берс, Д. М. Транспорт Na (+) в нормальном и больном сердце — помните о балансе. Журнал молекулярной и клеточной кардиологии 61, 2–10 (2013).
CAS PubMed PubMed Central Google Scholar
Tseng, D. Y. et al. Влияние станниокальцина 1 на поглощение кальция у эмбрионов рыбок данио (Danio rerio). Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология 296, R549–557 (2009).
CAS PubMed Google Scholar
Джексон, Л.Ф., Маккормик, С. Д., Мэдсен, С. С., Суонсон, П. и Салливан, К. В. Осморегуляторные эффекты гипофизэктомии и замены гомологичного пролактина у гибридных полосатых окуней. Comp Biochem Phys B 140, 211–218 (2005).
Google Scholar
Бревес, Дж. П., Маккормик, С. Д. и Карлстром, Р. О. Пролактин и костистые ионоциты: новые сведения о клеточных и молекулярных мишенях пролактина в эпителии позвоночных. Gen Comp Endocr 203, 21–28 (2014).
CAS PubMed Google Scholar
Schultheis, P.J. et al. Фенотип, напоминающий синдром Гительмана у мышей, лишенных апикального Na + -Cl- котранспортера дистального извитого канальца. Journal of Biological Chemistry 273, 29150–29155 (1998).
CAS PubMed Google Scholar
Nicolet-Barousse, L. et al. Инактивация гена ко-транспортера Na-Cl (NCC) связана с высокой МПК через почечный и костный механизмы: анализ пациентов с синдромом Гительмана и Ncc нулевых мышей.Журнал исследований костей и минералов: официальный журнал Американского общества исследований костей и минералов 20, 799–808 (2005).
CAS Google Scholar
Нгуен, Н., Стеллваг, Э. Дж. И Чжу, Ю. Пролактин-зависимая модуляция органогенеза у позвоночных: недавние открытия на рыбках данио. Comp Biochem Phys C 148, 370–380 (2008).
Google Scholar
Нгуен, Н.& Чжу, Ю. Пролактин функционирует как фактор выживания во время эмбриогенеза рыбок данио. Comp Biochem Phys A 153, 88–93 (2009).
Google Scholar
Эванс, Д. Х. Осморегуляция костистых рыб: чему мы научились со времен Августа Крога, Гомера Смита и Ансела Киз. Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология 295, R704–713 (2008).
CAS PubMed Google Scholar
Смит, Х.W. Регуляция воды и ее эволюция у рыб. Q Rev Biol 7, 1-26 (1932).
CAS Google Scholar
Hentschel, D. M. et al. Быстрый скрининг целостности щелевой диафрагмы клубочка у личинок рыбок данио. Am J Physiol-Renal 293, F1746 – F1750 (2007).
ADS CAS Google Scholar
Райдер, С.А. и др. Методы исследования in vivo сердечно-почечной функции у личинок рыбок данио (Danio rerio).J. Physiol-London 590, 1803–1809 (2012).
CAS PubMed Google Scholar
Cermak, T. et al. Эффективный дизайн и сборка пользовательских TALEN и других эффекторных конструкций на основе TAL для нацеливания на ДНК. Исследование нуклеиновых кислот 39, e82 (2011).
CAS PubMed PubMed Central Google Scholar
Moens, C. Цельная РНК in situ гибридизация на эмбрионах рыбок данио: гибридизация.Протоколы CSH 2008, pdb prot5037 (2008).
Moens, C. Цельная РНК in situ гибридизация на эмбрионах рыбок данио: сборка. Протоколы CSH 2008, pdb prot5038 (2008).
Moens, C. Цельная РНК in situ гибридизация на эмбрионах рыбок данио: синтез зонда. Протоколы CSH 2008, pdb prot5036 (2008).
Xu, Z. W. et al. Производство, характеристика и применение мышиных моноклональных антител против гонадотропина, соматолактина и пролактина из карпа обыкновенного (Cyprinus carpio).Gen Comp Endocr 167, 373–378 (2010).
CAS PubMed Google Scholar
Walker, M. B. & Kimmel, C. B. Двухцветное бескислотное окрашивание хрящей и костей для личинок рыбок данио. Биотехника и гистохимия: официальная публикация Комиссии по биологическим пятнам 82, 23–28 (2007).
CAS Google Scholar
Esaki, M. et al. Визуализация у личинок рыбок данио захвата Na + в богатых митохондриями клетках, дифференцировка которых зависит от foxi3a.AJP: Регуляторная, интегративная и сравнительная физиология 292, R470 – R480 (2006).
Google Scholar
Гиперпролактинемия (нарушение пролактации) | ColumbiaDoctors
Что такое гиперпролактинемия?
Гиперпролактинемия — это наличие аномально высокого уровня пролактина в крови. Пролактин — это гормон, вырабатываемый гипофизом. Многочисленные функции пролактина в организме в основном связаны с беременностью и производством грудного молока новорожденным.Однако уровень пролактина может повышаться, когда женщина не беременна или не кормит грудью, вызывая ряд состояний, которые могут повлиять на нормальную менструальную функцию и фертильность.
Что вызывает гиперпролактинемию?
Аномальное повышение уровня пролактина может происходить, когда продуцирующие пролактин клетки внутри гипофиза, называемые лактотрофами, производят больше гормона, чем требуется, или когда лактотрофы ненормально растут с образованием опухолей. Повышенный уровень пролактина также может возникать как побочный эффект некоторых психиатрических препаратов.
Как диагностируется гиперпролактинемия?
Простые анализы крови для измерения количества пролактина в крови могут подтвердить диагноз повышенного уровня пролактина. Уровень пролактина выше 25 нг / мл у небеременных женщин считается повышенным. Поскольку каждый человек испытывает ежедневные колебания уровня пролактина, может потребоваться повторить анализ крови, если уровень гормона повышен лишь незначительно. Многие женщины получают этот диагноз после обследования на бесплодие или нерегулярные периоды, но у других симптомы отсутствуют вообще.Иногда у пациентов появляются спонтанные молочные выделения из сосков, но у большинства этот симптом не проявляется. Умеренное повышение пролактина, от 25 до 50 нг / мл, обычно не вызывает заметных изменений менструального цикла, хотя может снизить общую фертильность. Более высокие уровни пролактина от 50 до 100 нг / мл могут вызвать нерегулярные менструальные периоды и значительно снизить фертильность женщины. Уровни пролактина более 100 нг / мл могут изменить нормальную функцию репродуктивной системы женщины, вызывая симптомы менопаузы (отсутствие менструаций, приливы, сухость влагалища) и бесплодие.
Как лечится гиперпролактинемия?
Лекарства, которые имитируют химический состав мозга дофамин, могут успешно использоваться для лечения большинства пациентов с повышенным уровнем пролактина. Эти лекарства ограничивают выработку пролактина гипофизом и вызывают регрессию пролактин-продуцирующих клеток. Два наиболее часто назначаемых лекарства — это каберголин (Достинекс®) и бромокриптин (Парлодел®). Начав с небольшой дозы, которую постепенно увеличивают, можно свести к минимуму побочные эффекты, в том числе изменения артериального давления и умственную «затуманенность».«Пациенты обычно хорошо реагируют на это лекарство, и уровень пролактина упадет через 2–3 недели. Как только уровень пролактина упадет, лекарство можно скорректировать для поддержания нормального уровня пролактина, а иногда можно полностью прекратить прием. Спонтанный регресс опухоли обычно происходит в течение нескольких лет без каких-либо клинических последствий.
У небольшого меньшинства пациентов лекарства неэффективны для снижения уровня пролактина, и большие опухоли (макроаденомы) сохраняются. Эти пациенты являются кандидатами на хирургическое лечение (резекция трансфеноидальной аденомы) и / или лучевая терапия.
Добавить комментарий